

Review
doi: 10.1038/bjc.2016.412.
Epub 2016 Dec 22.
Tumour microenvironment factors shaping the cancer metabolism landscape
Affiliations
- PMID: 28006817
- PMCID: PMC5294476
- DOI: 10.1038/bjc.2016.412
Item in Clipboard
Review
Tumour microenvironment factors shaping the cancer metabolism landscape
Br J Cancer.
2017 Jan.
Abstract
Cancer cells exhibit metabolic alterations that distinguish them from healthy tissues and make their metabolic processes susceptible to pharmacological targeting. Although typical cell-autonomous features of cancer metabolism have been emerging, it is increasingly appreciated that extrinsic factors also influence the metabolic properties of tumours. This review highlights evidence from the recent literature to discuss how conditions within the tumour microenvironment shape the metabolic character of tumours.
Conflict of interest statement
The author declares no conflict of interest.
Figures

Overview of pathways implicated in cancer metabolism. Key enzymes are shown in white boxes. See text for details. Coloured block arrows denote contribution of metabolites to the respective cellular processes. ΔΨm denotes mitochondrial inner membrane potential. -CH3 and -COCH3 denote methyl and acetyl groups, respectively. ALT, alanine aminotransferase; 1,3-BG, 1,3-bisphosphoglycerate; DHAP, dihydroxyacetone phosphate; F-1,6-BP, fructose-1, 6-bisphosphate; F-6-P, fructose-6-phosphate; FH, fumarate hydratase; G-3-P, glyceraldehyde-3-phosphate; G-6-P, glucose-6-phosphate; GLS, glutaminase, GS, glutamine synthetase; α-KG, α-ketoglutarate; HK, hexokinase; IDH, isocitrate dehydrogenase; LDH, lactate dehydrogenase; OXPHOS, oxidative phosphorylation; PC, pyruvate carboxylase; PDH, pyruvate dehydrogenase; PFK, phosphofructokinase; PKM, pyruvate kinase M isoform; PEP, phosphoenolpyruvate; 2-PG, 2-phosphoglycerate; 3-PG, 3-phosphoglycerate; ROS, reactive oxygen species; SDH, succinate dehydrogenase.
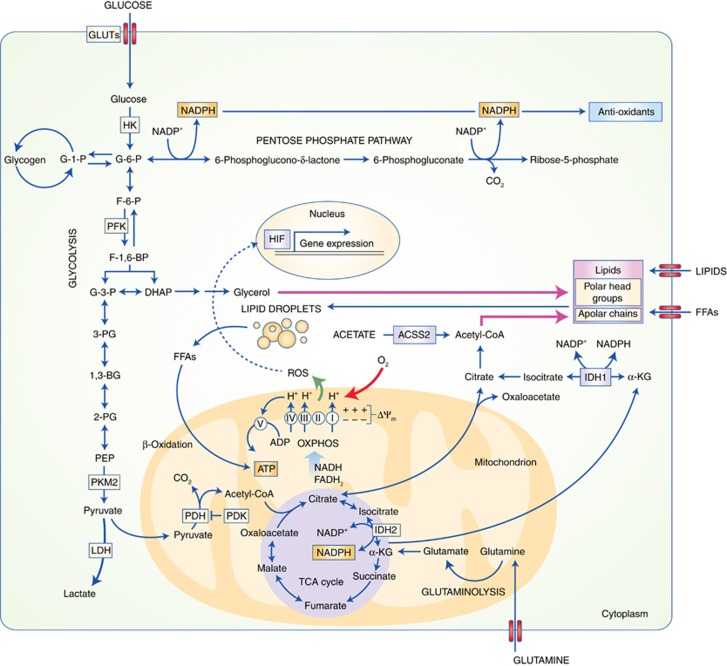
Overview of key metabolic pathways implicated in the response of cancer cells to low oxygen conditions. See text for details. ACSS2, Acetyl-CoA synthase isoform 2; HIF, hypoxia-inducible factor; FFAs, free fatty acids; other abbreviations are as in Figure 1.
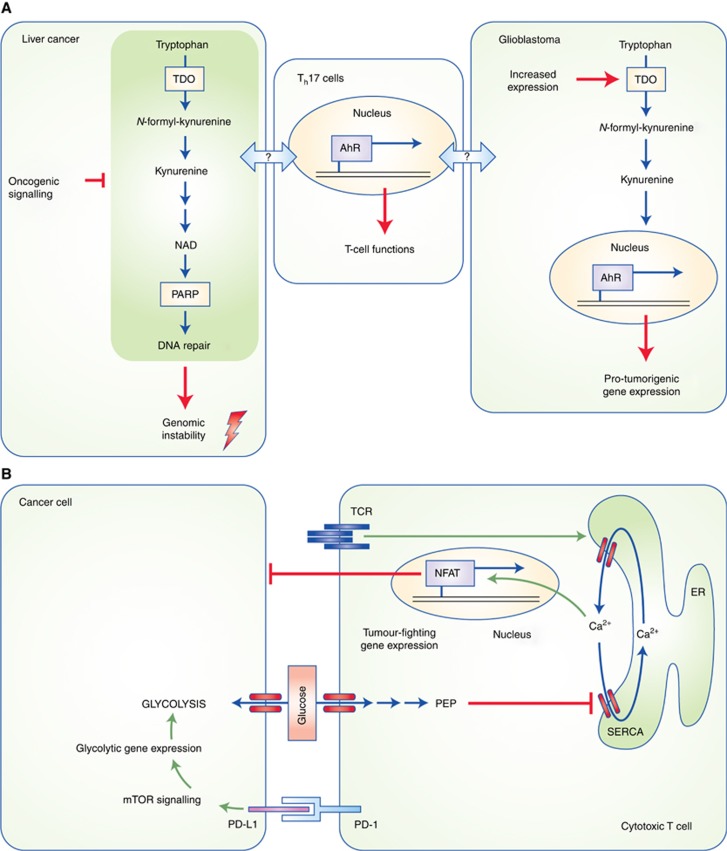
Paradigms of cancer–immune cell interactions. (A) Tryptophan metabolism yields the intermediate kynurenine, which promotes glioblastoma development by activating the aryl hydrocarbon receptor, a transcription factor that controls the expression of pro-tumorigenic genes. A downstream product of further kynurenine catabolism is NAD, which, among others, is an essential co-factor for the DNA repair enzyme poly-ADP ribose polymerase (PARP). Suppression of NAD synthesis by oncogenic signalling in liver tumours leads to increased genomic instability, thereby promoting tumorigenesis. Th17 cell differentiation can be influenced by tryptophan metabolites and Th17-mediated inflammation has been implicated in liver cancer. Whether tryptophan metabolism in cancer cells influences tumour progression by modulating immune cells remains to be seen. (B) Metabolic competition for glucose between cancer cells and tumour-infiltrating T cells. In the proximity of highly glycolytic cancer cells, T cells fail to sustain cytoplasmic calcium levels necessary to activate the NFAT transcription factor, which mediates anti-tumour responses of T cells. The immunosuppressive molecule PD-L1 in cancer cells, may further promote glucose metabolism by signalling through the mechanistic target of rapamycin (mTOR) pathway, in addition to suppressing T cells via interaction with PD-1. ER, endoplasmic reticulum; NFAT, nuclear factor of activated T cells; PD-1, programmed cell death protein 1; PD-L1, PD ligand 1; PEP; phosphoenolpyruvate; SERCA, sarcoplasmic/endoplasmic reticulum calcium ATPase; TCR, T-cell receptor; TDO, tryptophan dioxygenase.
References
-
- Anastasiou D, Poulogiannis G, Asara JM, Boxer MB, Jiang JK, Shen M, Bellinger G, Sasaki AT, Locasale JW, Auld DS, Thomas CJ, Vander Heiden MG, Cantley LC (2011) Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 334: 1278–1283. - PMC - PubMed
-
- Bensaad K, Favaro E, Lewis CA, Peck B, Lord S, Collins JM, Pinnick KE, Wigfield S, Buffa FM, Li JL, Zhang Q, Wakelam MJ, Karpe F, Schulze A, Harris AL (2014) Fatty acid uptake and lipid storage induced by HIF-1alpha contribute to cell growth and survival after hypoxia-reoxygenation. Cell Rep 9: 349–365. - PubMed
-
- Bhutia YD, Babu E, Ramachandran S, Ganapathy V (2015) Amino Acid transporters in cancer and their relevance to ‘glutamine addiction’: novel targets for the design of a new class of anticancer drugs. Cancer Res 75: 1782–1788. - PubMed
-
- Buescher JM, Antoniewicz MR, Boros LG, Burgess SC, Brunengraber H, Clish CB, DeBerardinis RJ, Feron O, Frezza C, Ghesquiere B, Gottlieb E, Hiller K, Jones RG, Kamphorst JJ, Kibbey RG, Kimmelman AC, Locasale JW, Lunt SY, Maddocks OD, Malloy C, Metallo CM, Meuillet EJ, Munger J, Noh K, Rabinowitz JD, Ralser M, Sauer U, Stephanopoulos G, St-Pierre J, Tennant DA, Wittmann C, Vander Heiden MG, Vazquez A, Vousden K, Young JD, Zamboni N, Fendt SM (2015) A roadmap for interpreting (13)C metabolite labeling patterns from cells. Curr Opin Biotechnol 34: 189–201. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
