

The tumour microenvironment harbours ontogenically distinct dendritic cell populations with opposing effects on tumour immunity
- PMID: 28008905
- PMCID: PMC5196231
- DOI: 10.1038/ncomms13720
The tumour microenvironment harbours ontogenically distinct dendritic cell populations with opposing effects on tumour immunity
Abstract
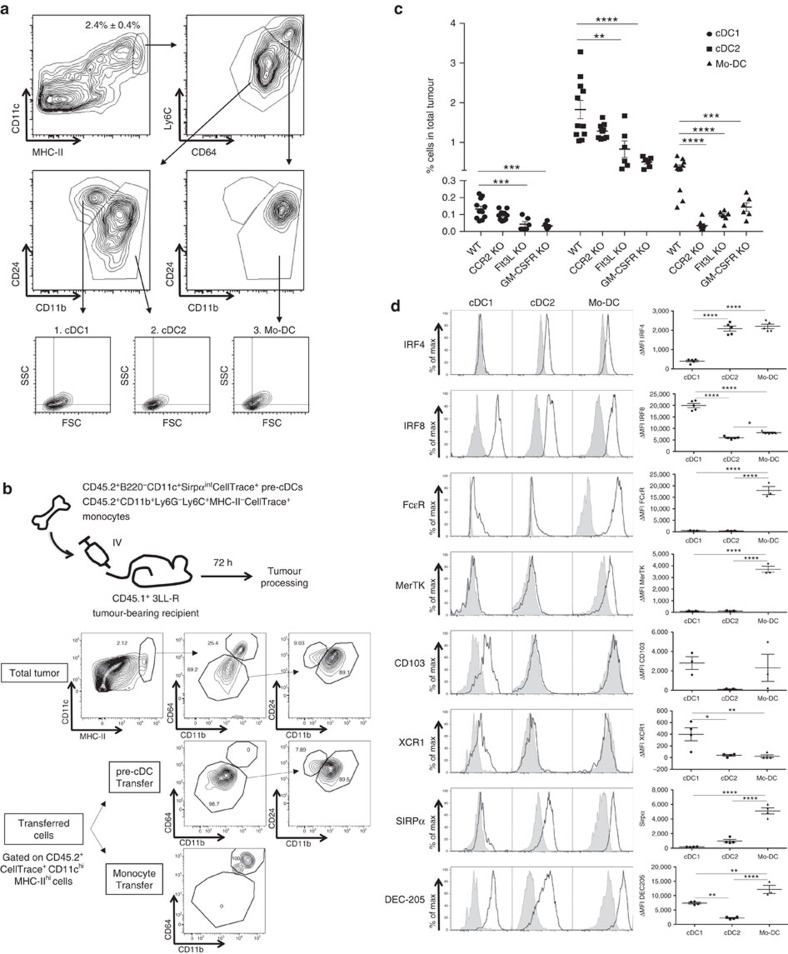
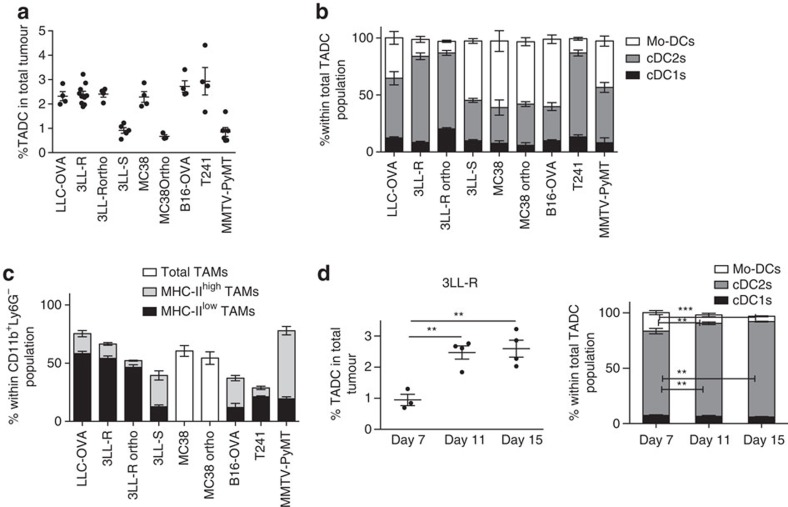
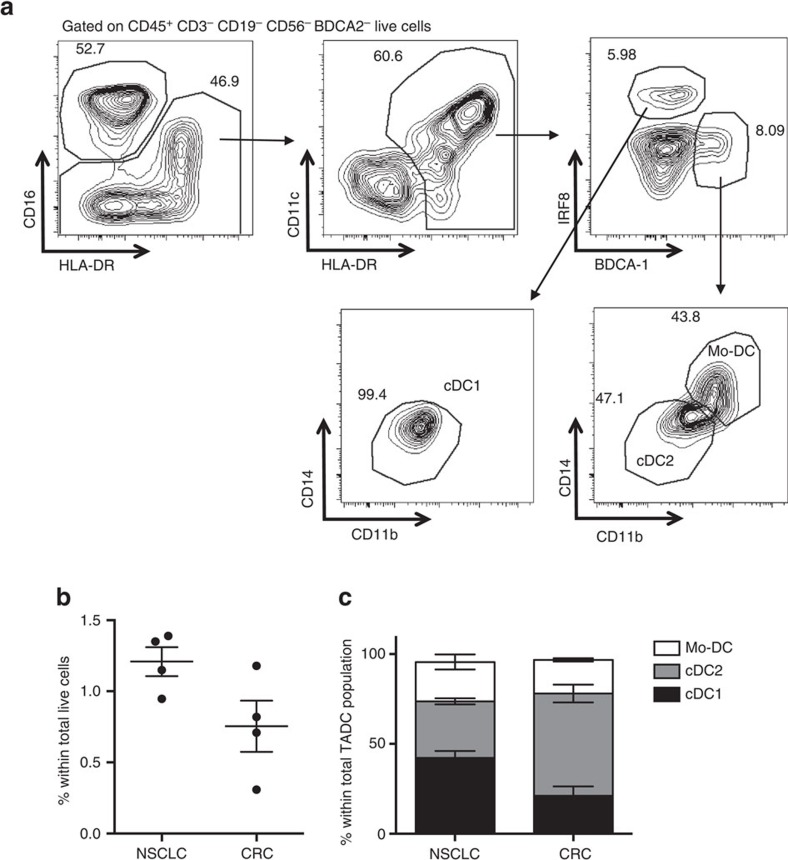
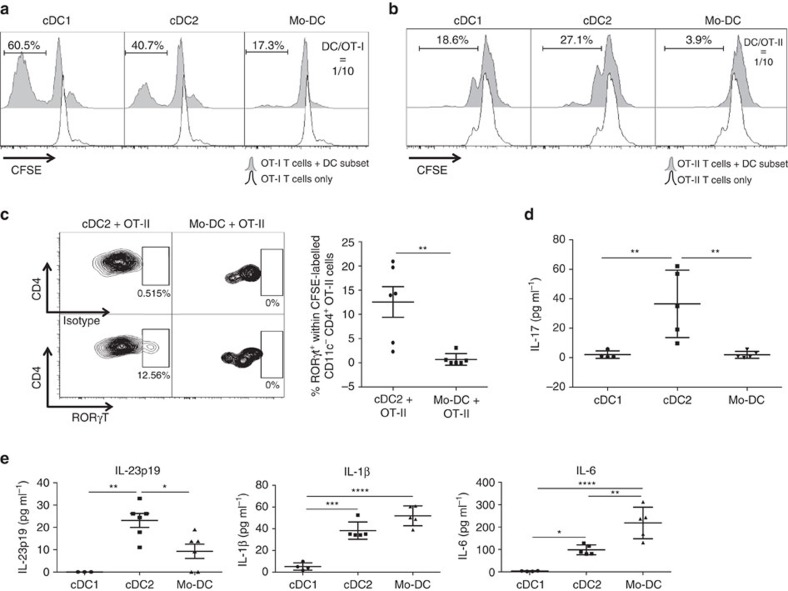
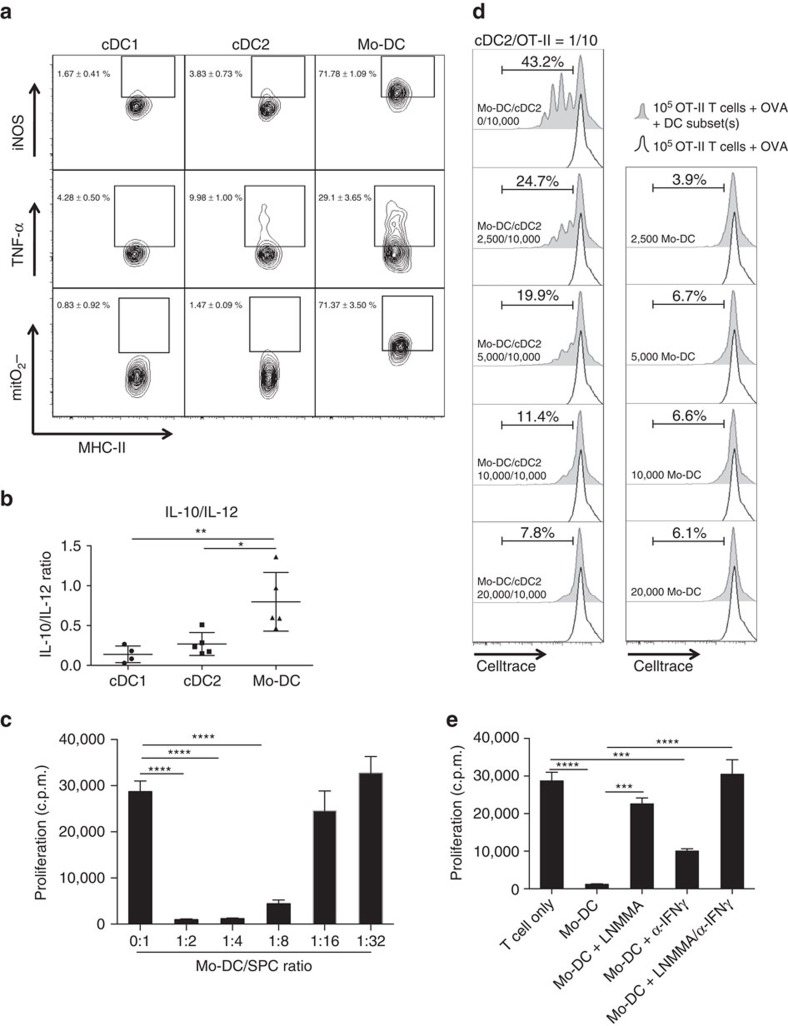
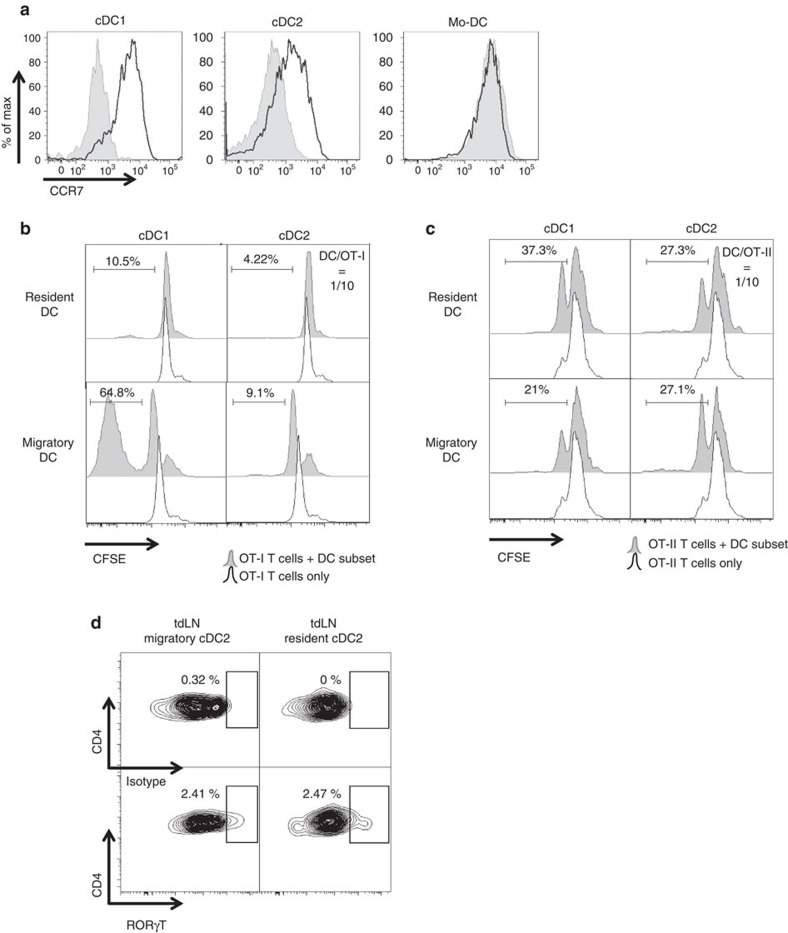
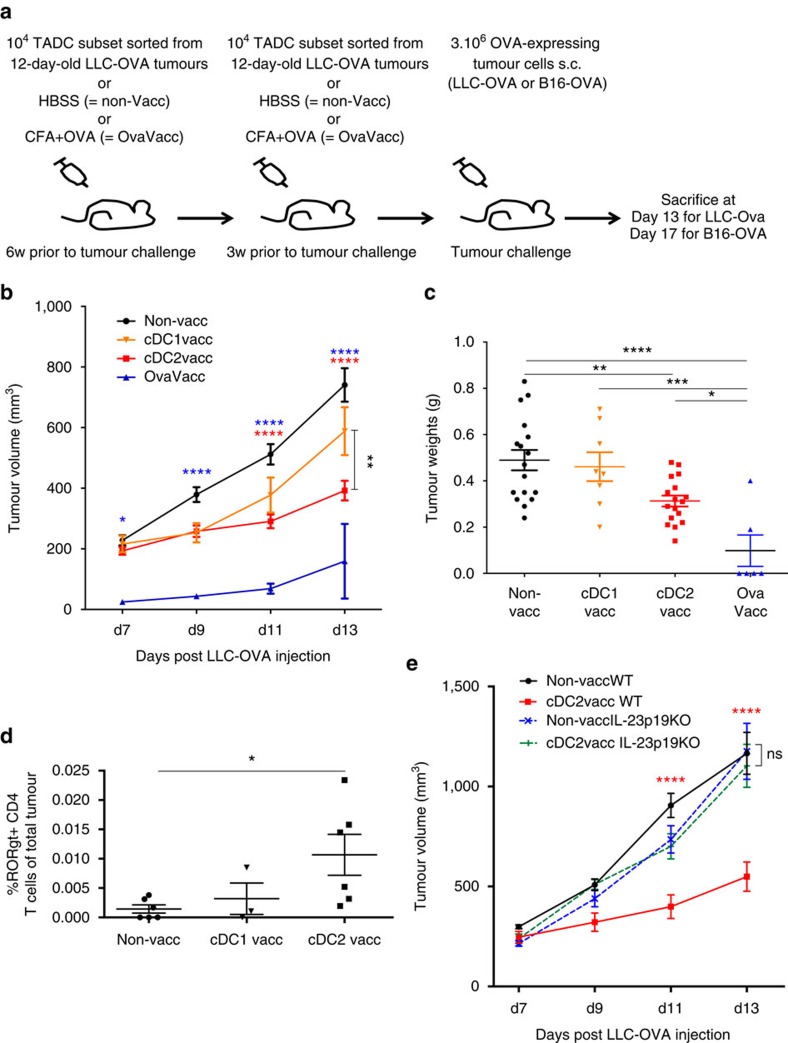
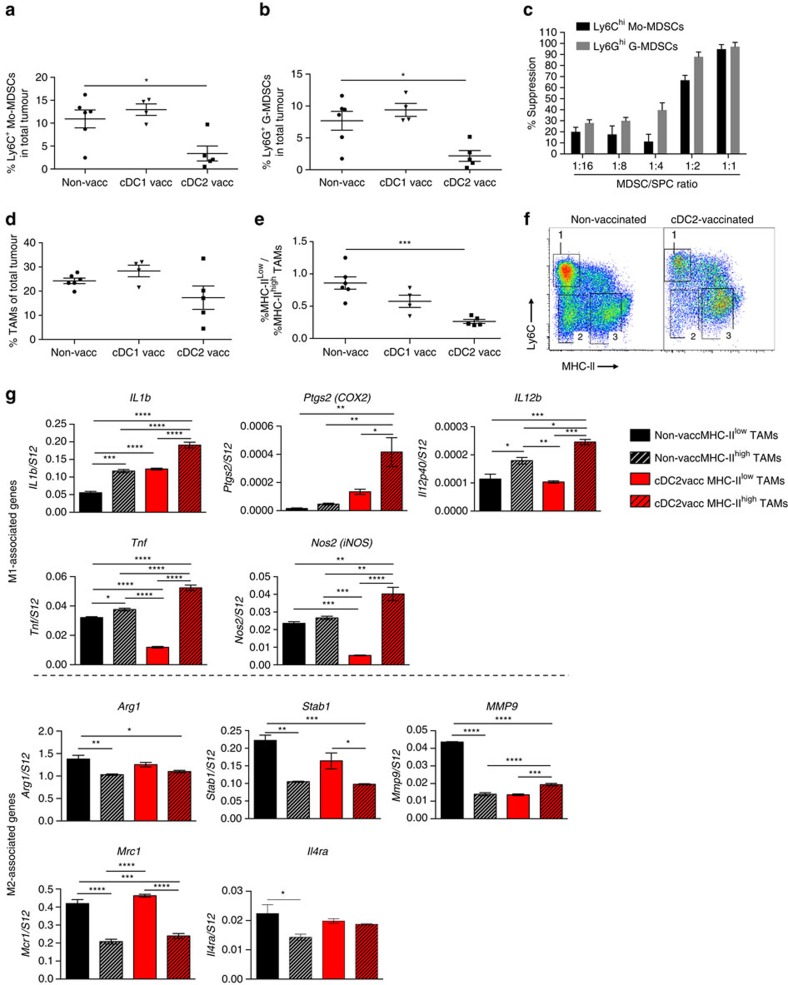
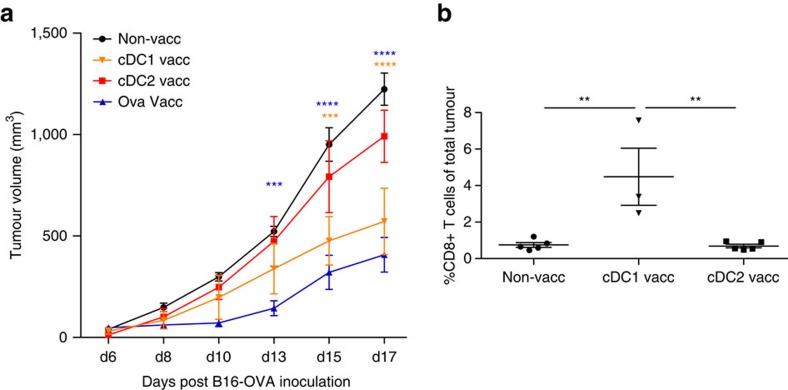
Various steady state and inflamed tissues have been shown to contain a heterogeneous DC population consisting of developmentally distinct subsets, including cDC1s, cDC2s and monocyte-derived DCs, displaying differential functional specializations. The identification of functionally distinct tumour-associated DC (TADC) subpopulations could prove essential for the understanding of basic TADC biology and for envisaging targeted immunotherapies. We demonstrate that multiple mouse tumours as well as human tumours harbour ontogenically discrete TADC subsets. Monocyte-derived TADCs are prominent in tumour antigen uptake, but lack strong T-cell stimulatory capacity due to NO-mediated immunosuppression. Pre-cDC-derived TADCs have lymph node migratory potential, whereby cDC1s efficiently activate CD8+ T cells and cDC2s induce Th17 cells. Mice vaccinated with cDC2s displayed a reduced tumour growth accompanied by a reprogramming of pro-tumoural TAMs and a reduction of MDSCs, while cDC1 vaccination strongly induces anti-tumour CTLs. Our data might prove important for therapeutic interventions targeted at specific TADC subsets or their precursors.
Figures

References
-
- Steinman R. M. & Banchereau J. Taking dendritic cells into medicine. Nature 449, 419–426 (2007). - PubMed
-
- Goc J. et al. Dendritic cells in tumor-associated tertiary lymphoid structures signal a Th1 cytotoxic immune contexture and license the positive prognostic value of infiltrating CD8+ T cells. Cancer Res. 74, 705–715 (2014). - PubMed
-
- Fridman W. H. et al. Prognostic and predictive impact of intra- and peritumoral immune infiltrates. Cancer Res. 71, 5601–5605 (2011). - PubMed
-
- Gabrilovich D. Mechanisms and functional significance of tumour-induced dendritic-cell defects. Nat. Rev. Immunol. 4, 941–952 (2004). - PubMed
-
- Vicari A. P., Caux C. & Trinchieri G. Tumour escape from immune surveillance through dendritic cell inactivation. Semin Cancer Biol. 12, 33–42 (2002). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
