

A Brief History of Research on Mitotic Mechanisms
- PMID: 28009830
- PMCID: PMC5192435
- DOI: 10.3390/biology5040055
A Brief History of Research on Mitotic Mechanisms
Abstract
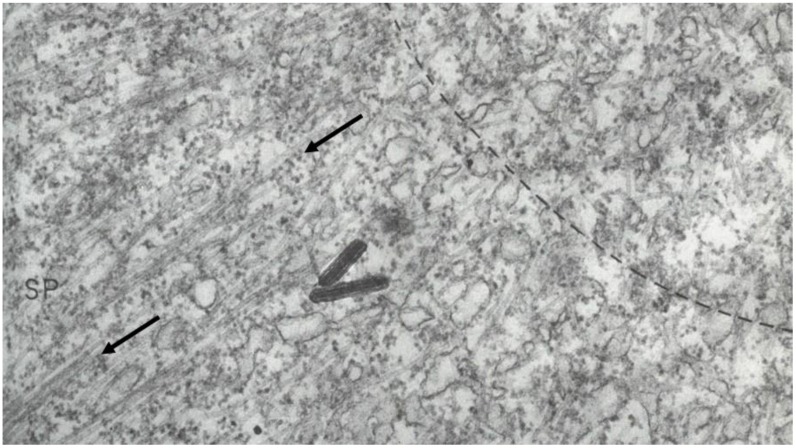
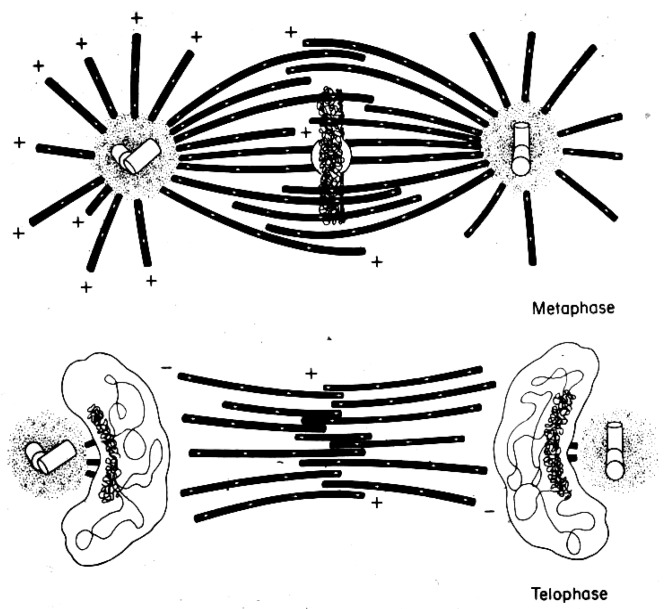
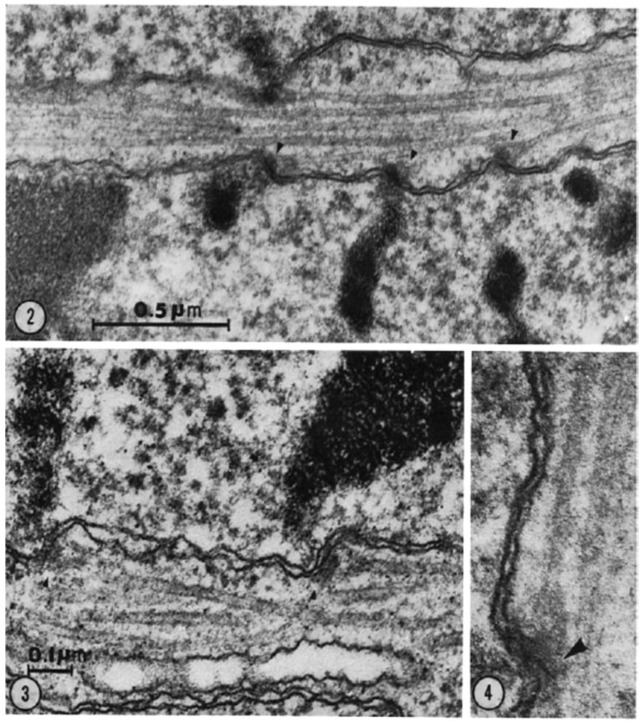
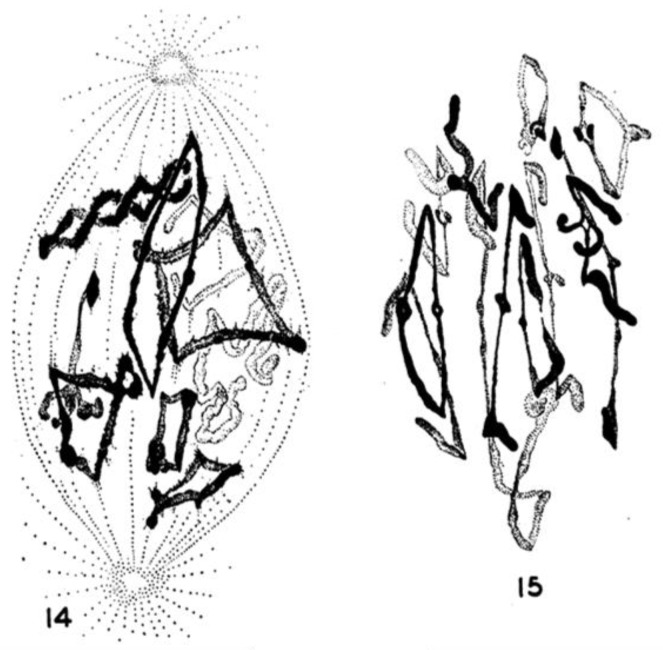
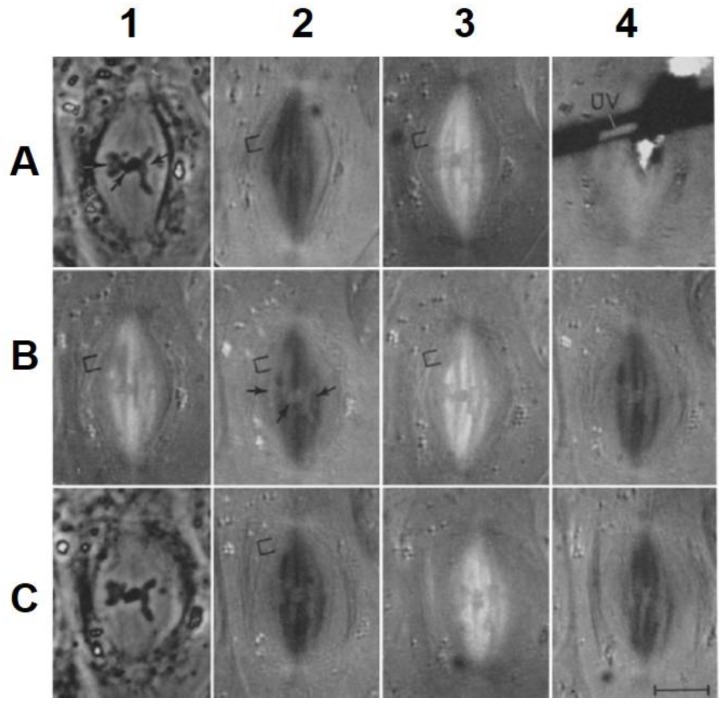
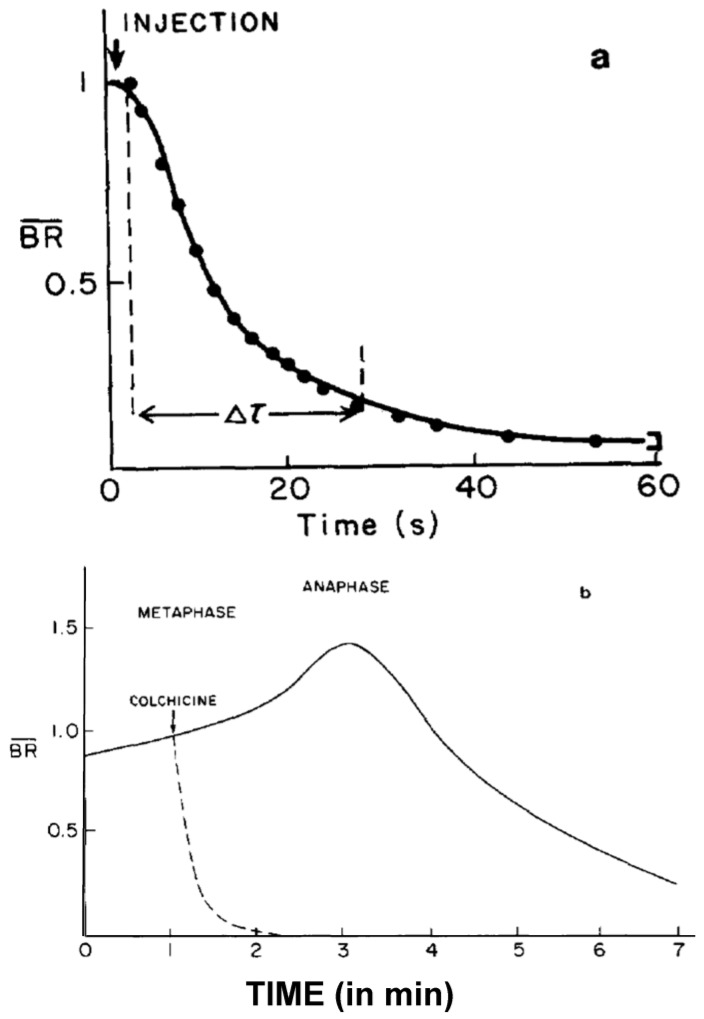
This chapter describes in summary form some of the most important research on chromosome segregation, from the discovery and naming of mitosis in the nineteenth century until around 1990. It gives both historical and scientific background for the nine chapters that follow, each of which provides an up-to-date review of a specific aspect of mitotic mechanism. Here, we trace the fruits of each new technology that allowed a deeper understanding of mitosis and its underlying mechanisms. We describe how light microscopy, including phase, polarization, and fluorescence optics, provided descriptive information about mitotic events and also enabled important experimentation on mitotic functions, such as the dynamics of spindle fibers and the forces generated for chromosome movement. We describe studies by electron microscopy, including quantitative work with serial section reconstructions. We review early results from spindle biochemistry and genetics, coupled to molecular biology, as these methods allowed scholars to identify key molecular components of mitotic mechanisms. We also review hypotheses about mitotic mechanisms whose testing led to a deeper understanding of this fundamental biological event. Our goal is to provide modern scientists with an appreciation of the work that has laid the foundations for their current work and interests.
Keywords: accuracy; centrosome; chromosome; force; kinetochore; microtubule; mitosis; mitotic spindle; motor enzyme; tubulin dynamics.
Conflict of interest statement
The authors state that they have no conflict of interests.
Figures



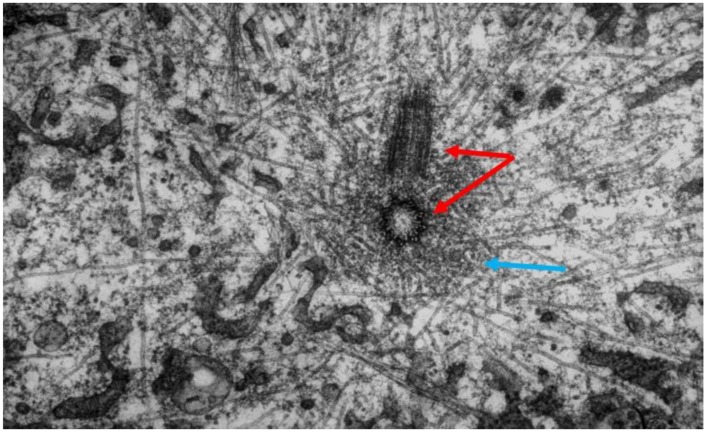

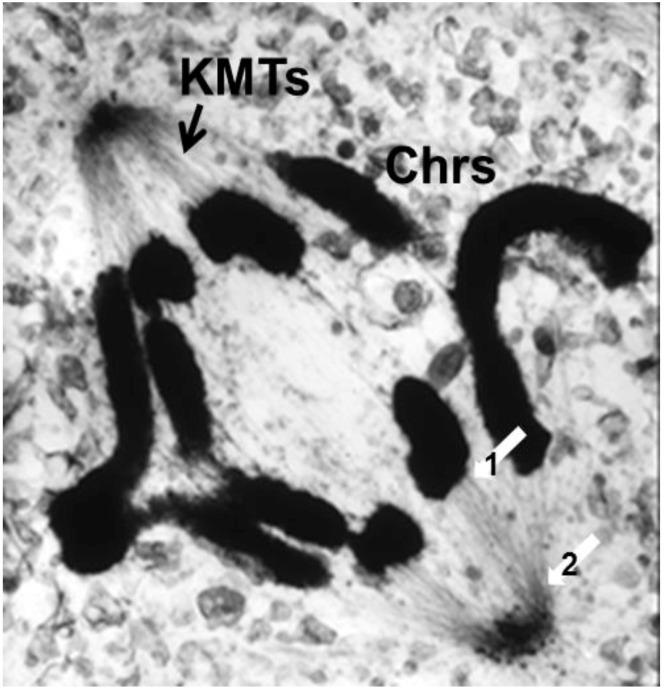
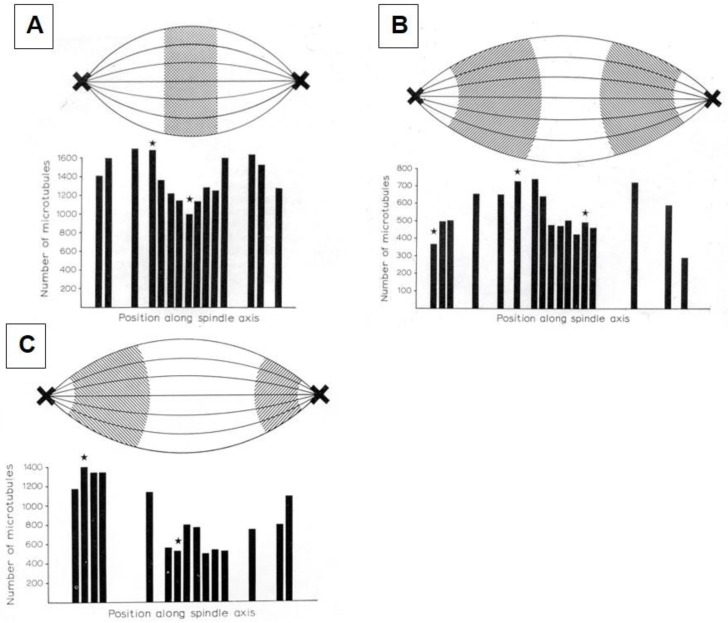
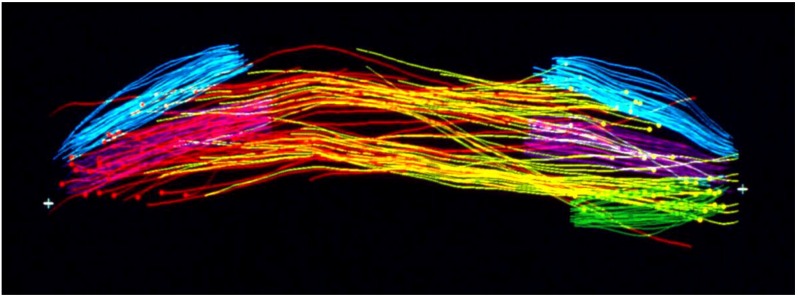




References
-
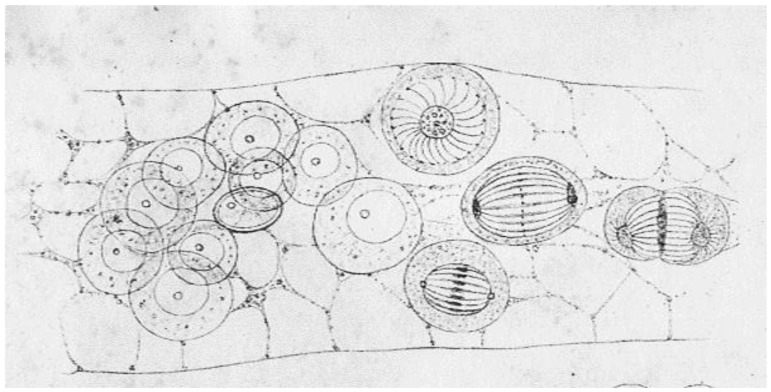
- Schneider A. Bericht der Oberhessischen Gesellschaft für Natur- und Heilkunde. Volume 14. Upper-Hessian Society for Natural and Medical Science; Giessen, Hesse, Germany: 1873. Untersuchungen über Plathelminthen. (In German)
-
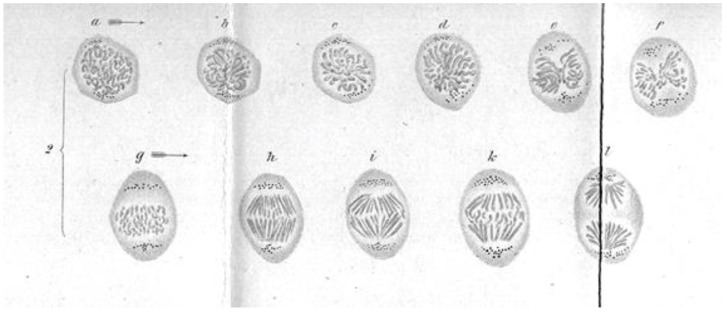
- Strasburger E. Zellbildung und Zelltheilung “Cell Formation and Cell Division”. Gustav Fischer; Jena, Germany: 1880.
-
- Van Beneden E. Recherches sur les Dicyemides. Bull. Acad. R. 1876;41:1–111.
-
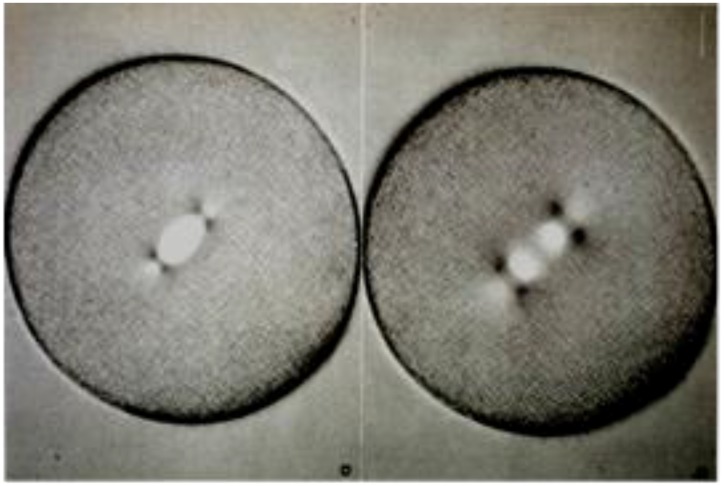
- Flemming W. Zur Kenntniss der Zelle und ihre Lebenserscheinungen. Arch. Mikr. Anat. 1878;16:302–436. doi: 10.1007/BF02956386. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
