

Suppression and synthetic-lethal genetic relationships of ΔgpsB mutations indicate that GpsB mediates protein phosphorylation and penicillin-binding protein interactions in Streptococcus pneumoniae D39
- PMID: 28010038
- PMCID: PMC5344783
- DOI: 10.1111/mmi.13613
Suppression and synthetic-lethal genetic relationships of ΔgpsB mutations indicate that GpsB mediates protein phosphorylation and penicillin-binding protein interactions in Streptococcus pneumoniae D39
Abstract
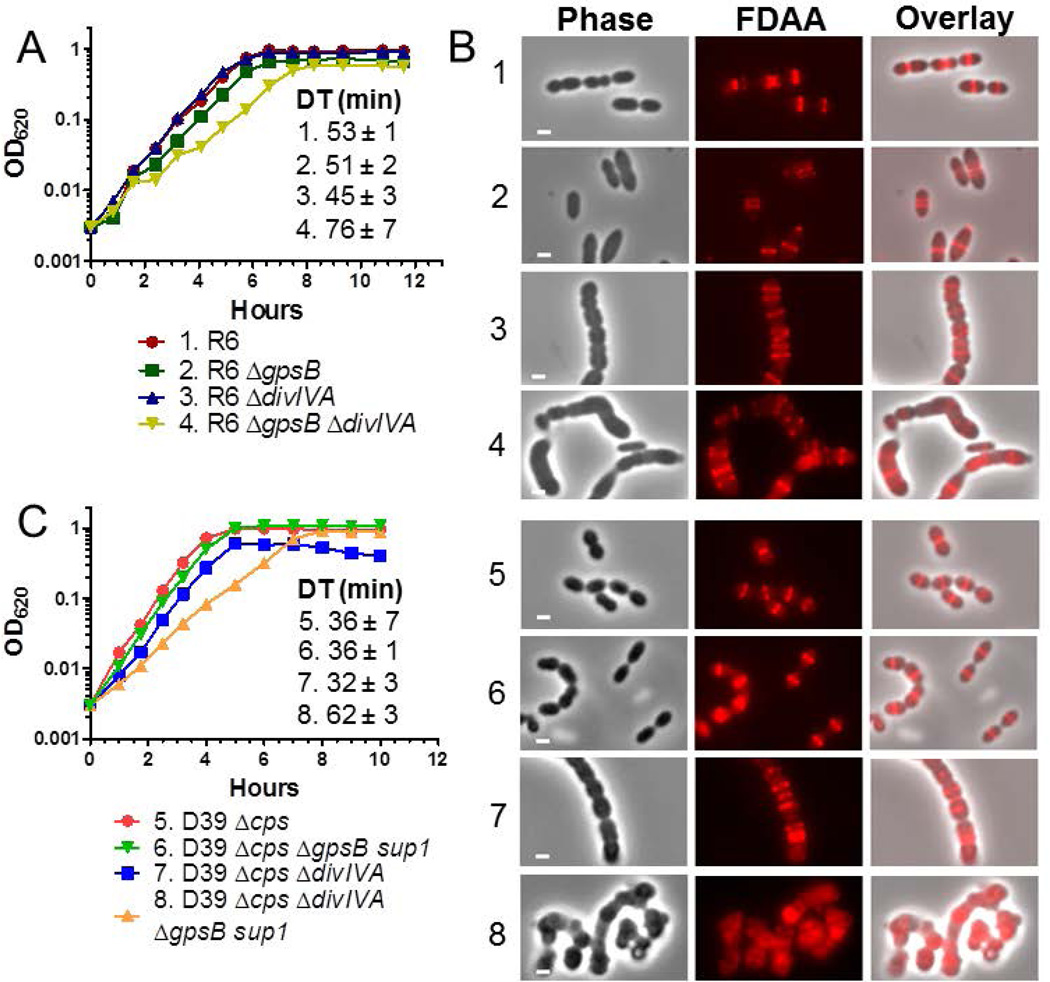
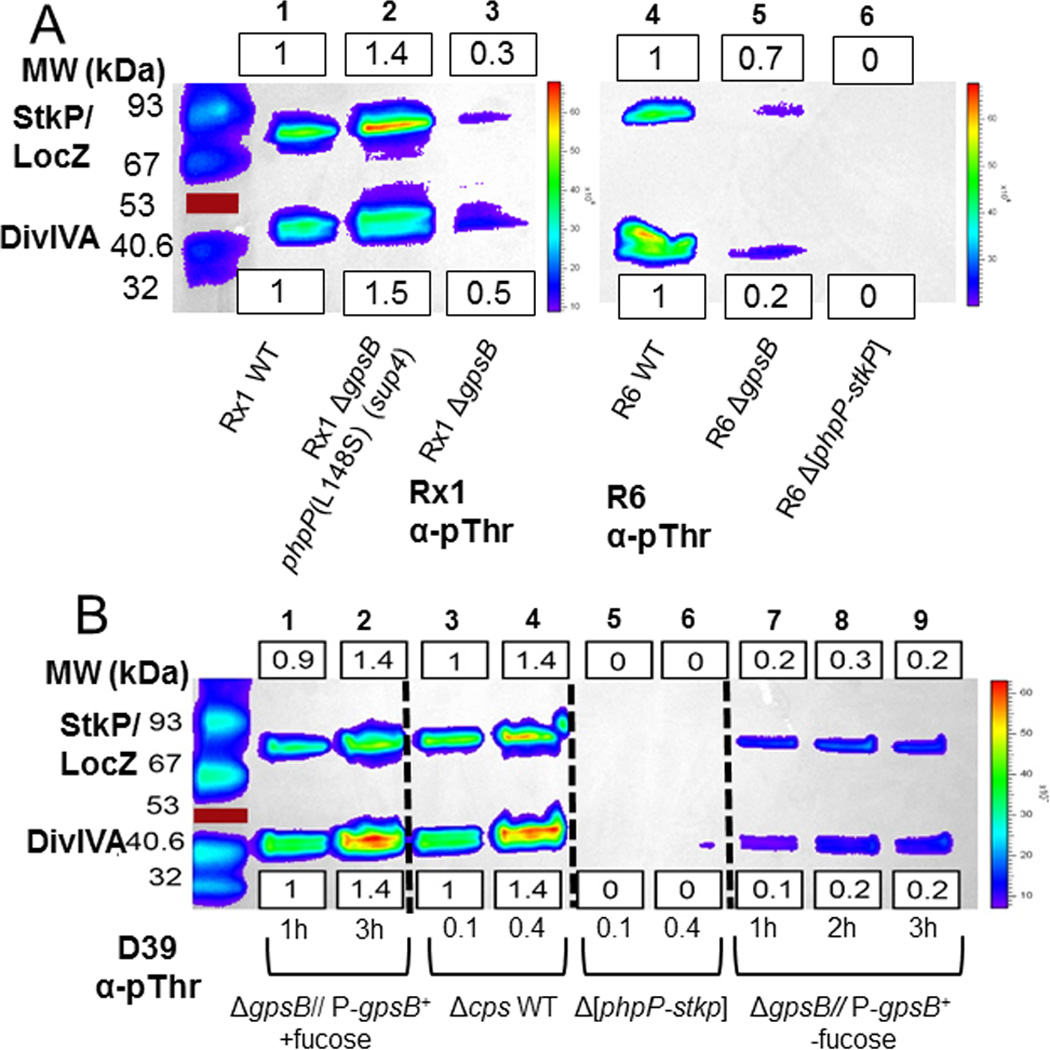
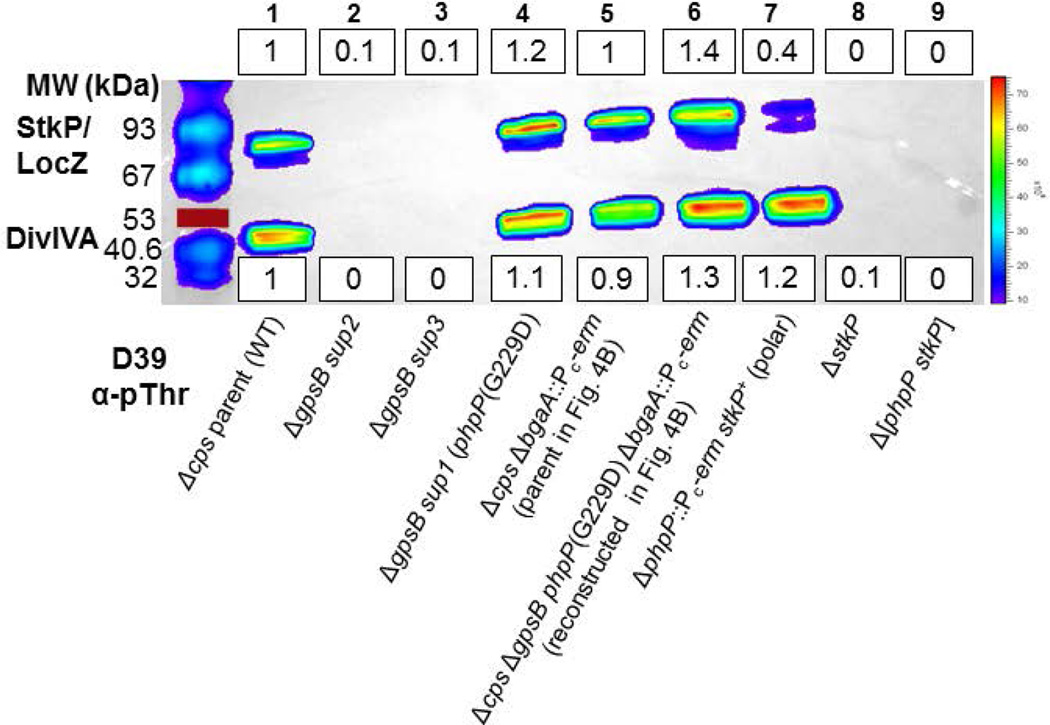
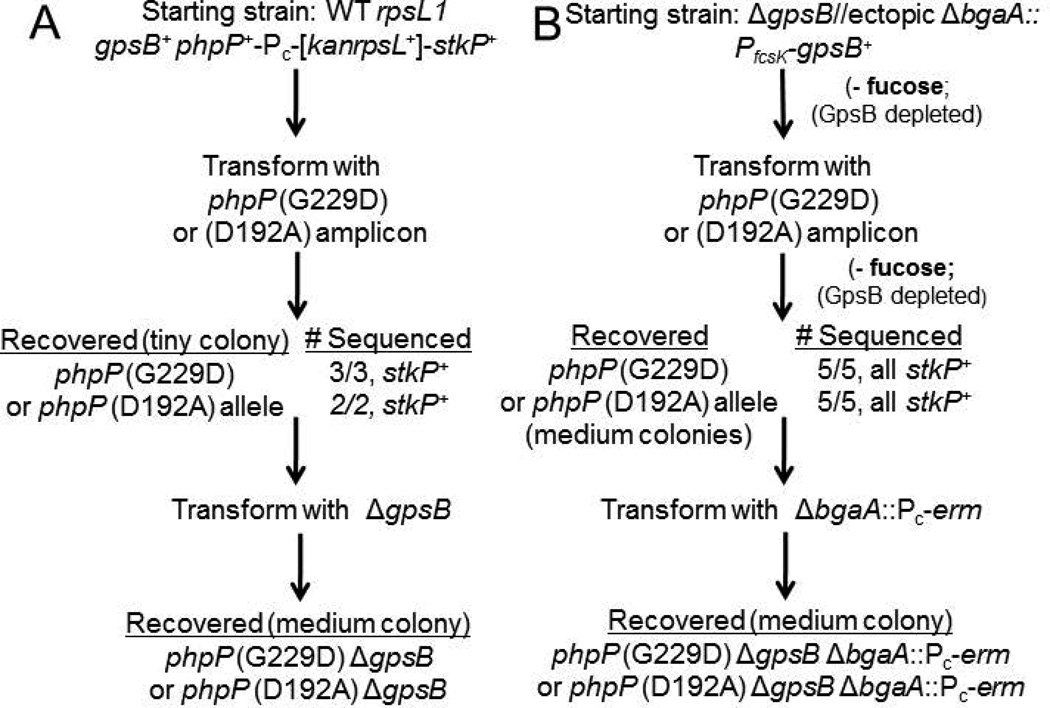
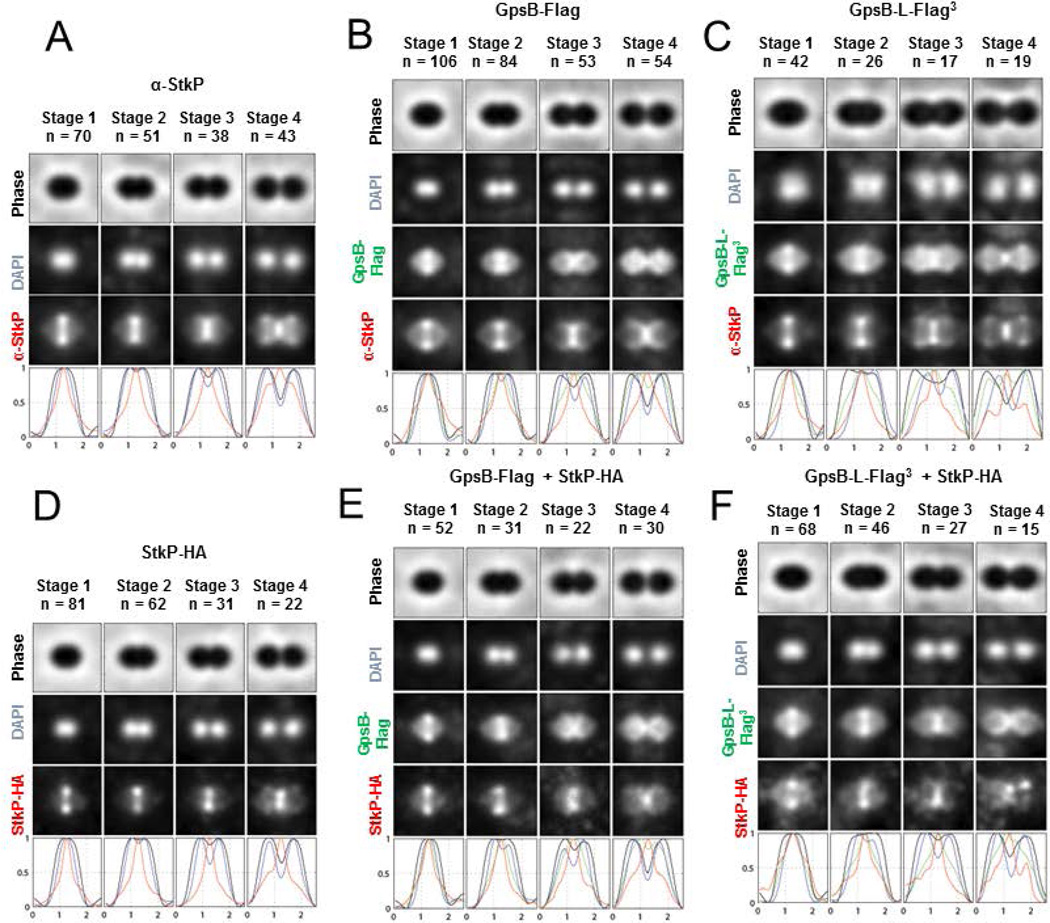
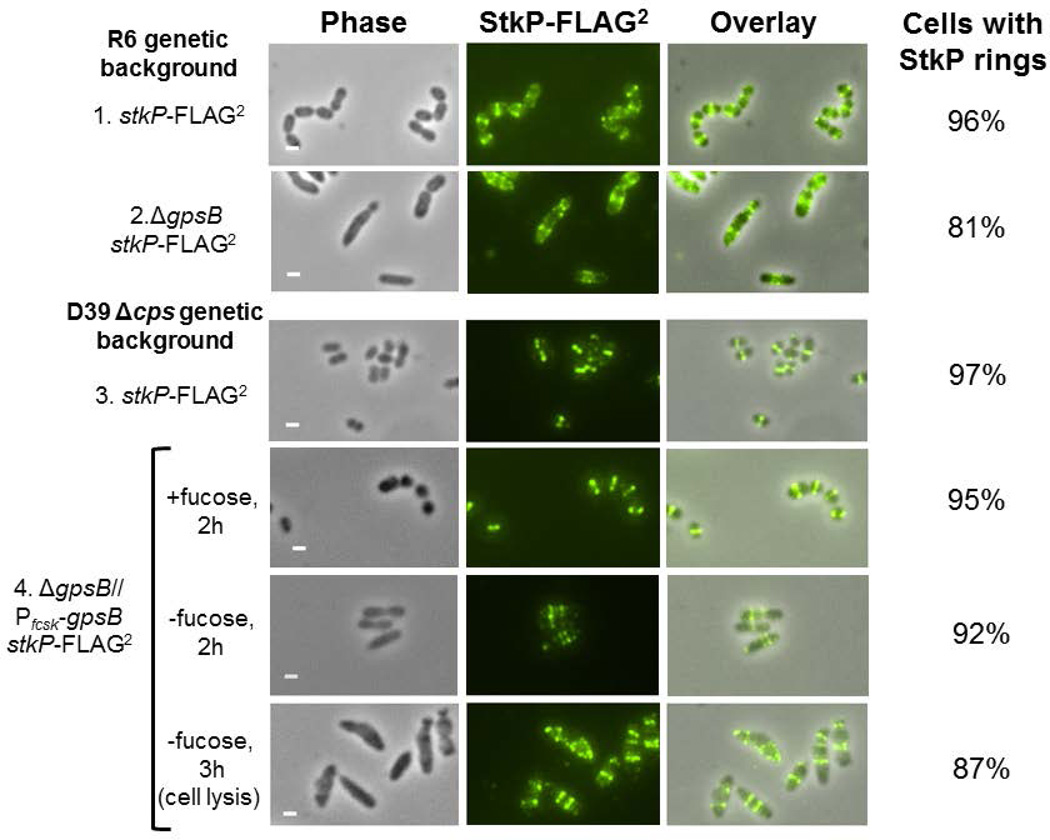
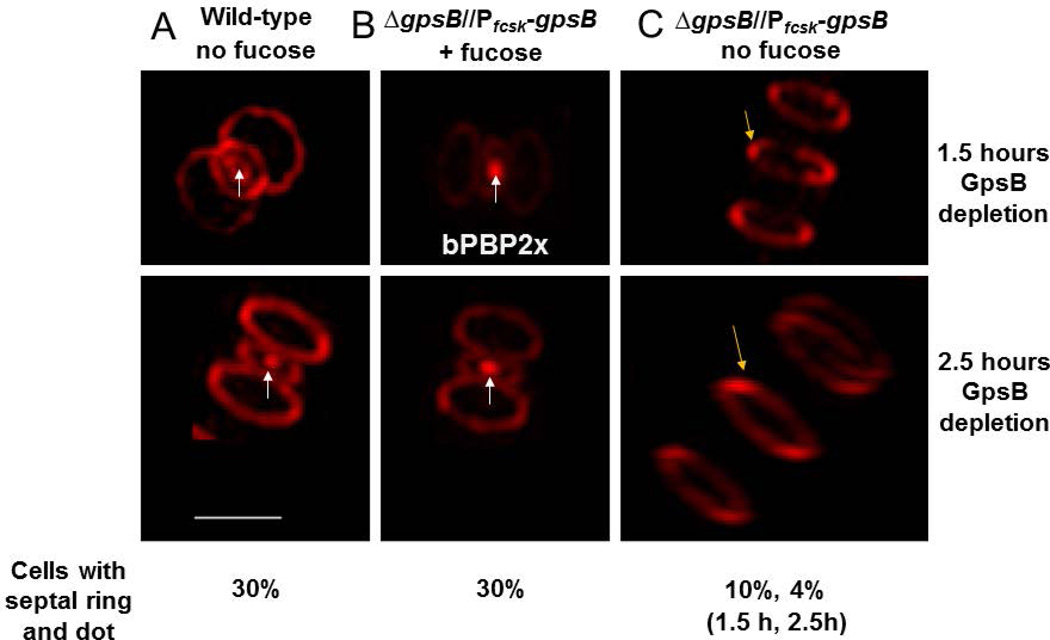
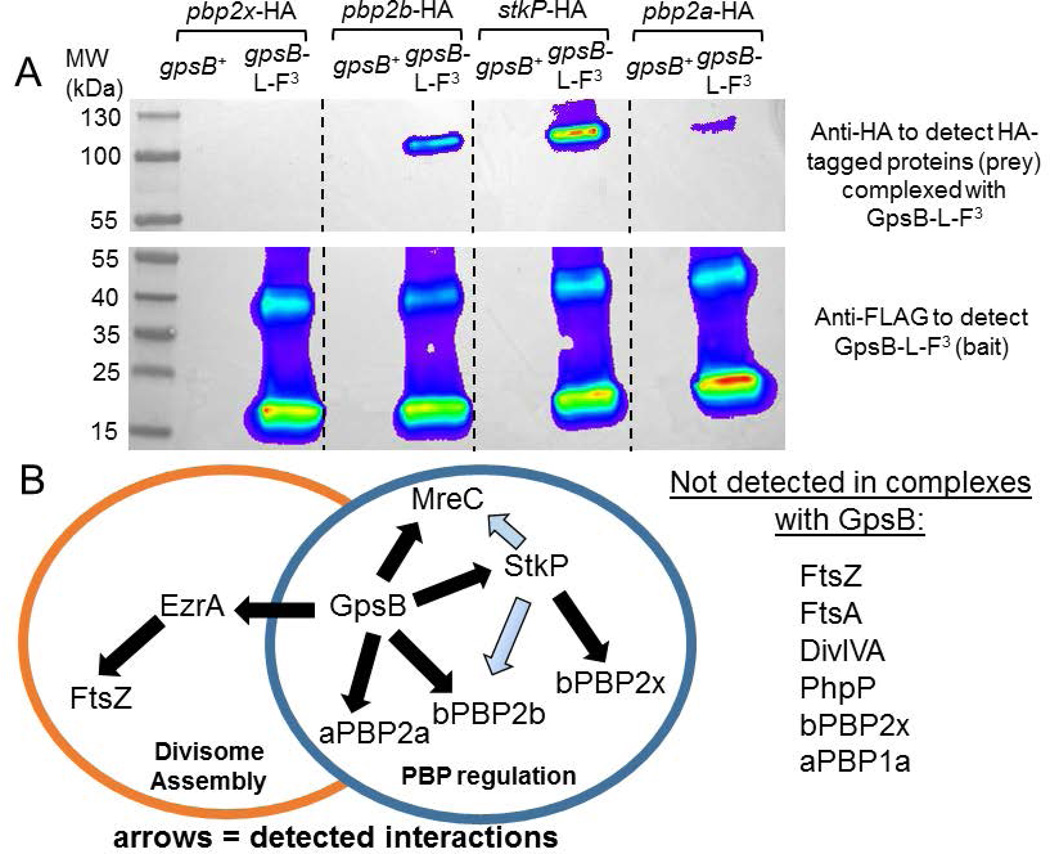
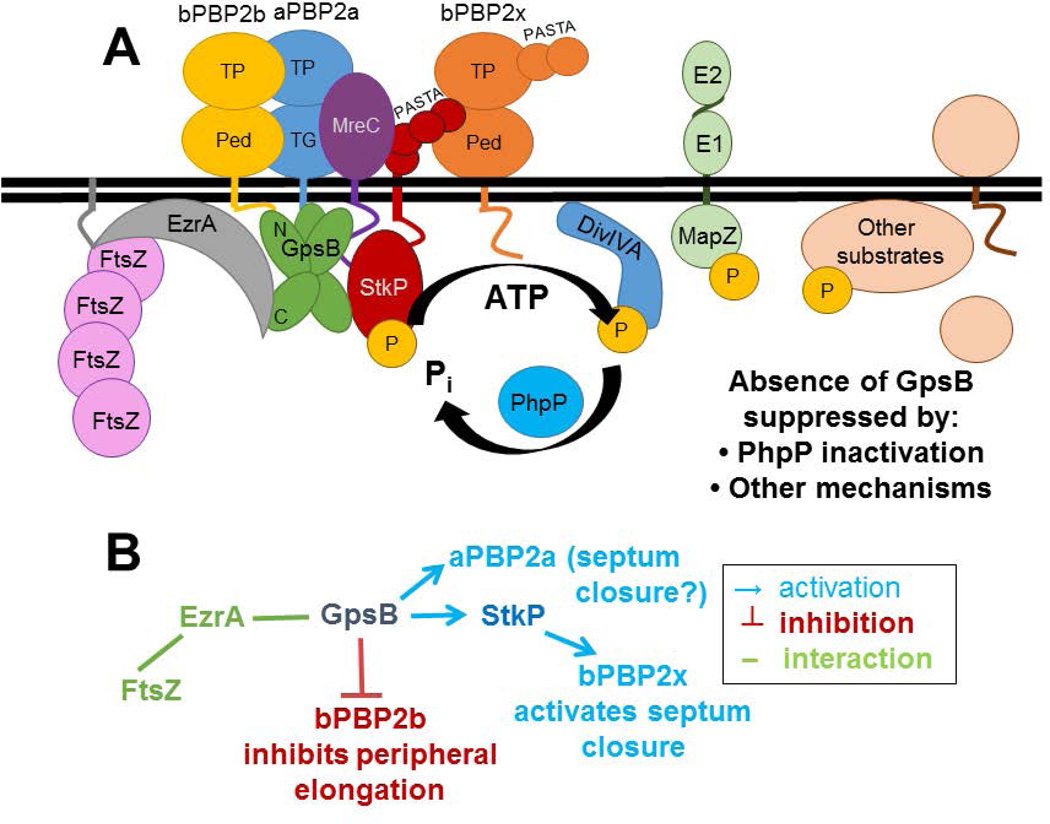
GpsB regulatory protein and StkP protein kinase have been proposed as molecular switches that balance septal and peripheral (side-wall like) peptidoglycan (PG) synthesis in Streptococcus pneumoniae (pneumococcus); yet, mechanisms of this switching remain unknown. We report that ΔdivIVA mutations are not epistatic to ΔgpsB division-protein mutations in progenitor D39 and related genetic backgrounds; nor is GpsB required for StkP localization or FDAA labeling at septal division rings. However, we confirm that reduction of GpsB amount leads to decreased protein phosphorylation by StkP and report that the essentiality of ΔgpsB mutations is suppressed by inactivation of PhpP protein phosphatase, which concomitantly restores protein phosphorylation levels. ΔgpsB mutations are also suppressed by other classes of mutations, including one that eliminates protein phosphorylation and may alter division. Moreover, ΔgpsB mutations are synthetically lethal with Δpbp1a, but not Δpbp2a or Δpbp1b mutations, suggesting GpsB activation of PBP2a activity. Consistent with this result, co-IP experiments showed that GpsB complexes with EzrA, StkP, PBP2a, PBP2b and MreC in pneumococcal cells. Furthermore, depletion of GpsB prevents PBP2x migration to septal centers. These results support a model in which GpsB negatively regulates peripheral PG synthesis by PBP2b and positively regulates septal ring closure through its interactions with StkP-PBP2x.
© 2016 John Wiley & Sons Ltd.
Figures

Comment in
-
The GpsB files: the truth is out there.Mol Microbiol. 2017 Mar;103(6):913-918. doi: 10.1111/mmi.13612. Epub 2017 Jan 12. Mol Microbiol. 2017. PMID: 28010044
References
-
- Archambaud C, Gouin E, Pizarro-Cerda J, Cossart P, Dussurget O. Translation elongation factor EF-Tu is a target for Stp, a serine-threonine phosphatase involved in virulence of Listeria monocytogenes. Molec Microbiol. 2005;56:383–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
