

Cell autonomous ANTXR1-mediated regulation of extracellular matrix components in primary fibroblasts
- PMID: 28011198
- PMCID: PMC5478475
- DOI: 10.1016/j.matbio.2016.12.002
Cell autonomous ANTXR1-mediated regulation of extracellular matrix components in primary fibroblasts
Abstract
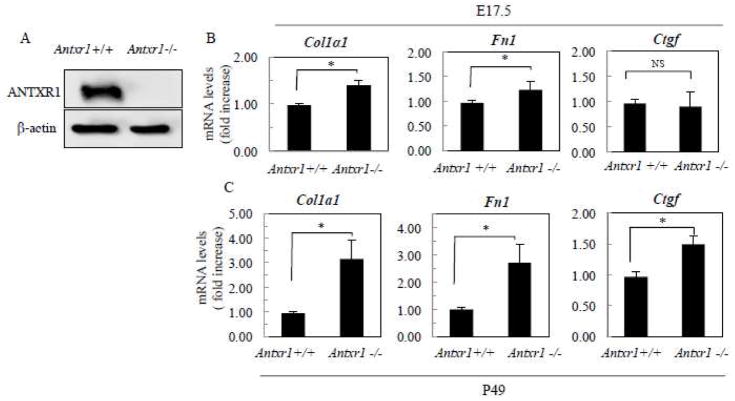
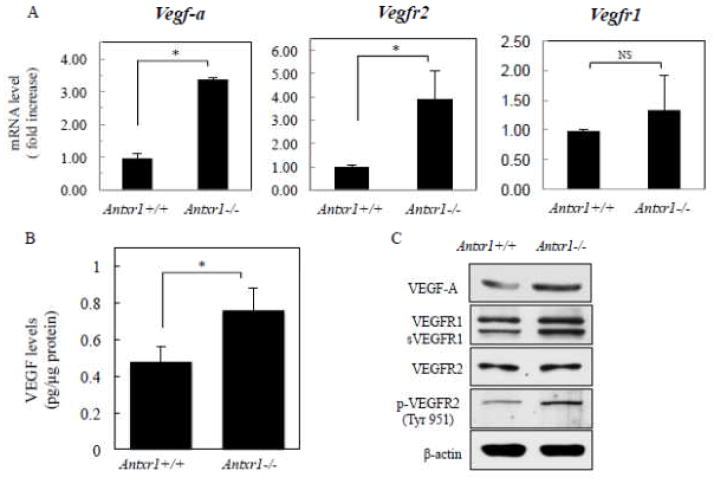
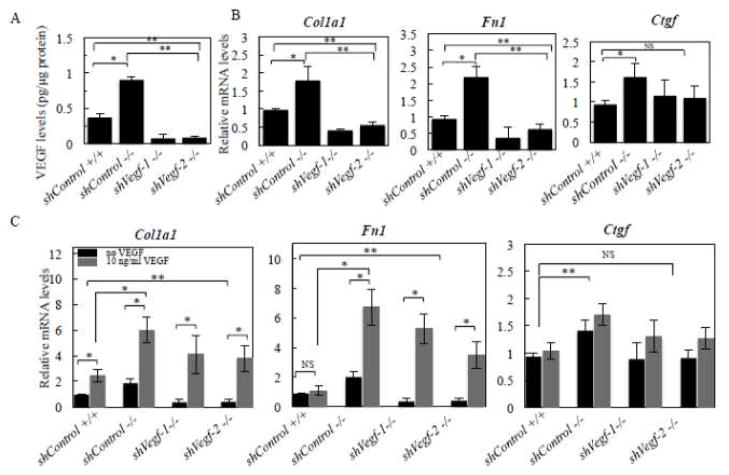
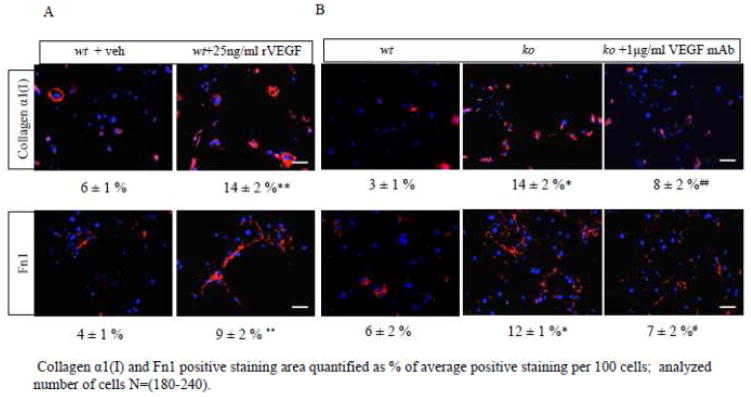
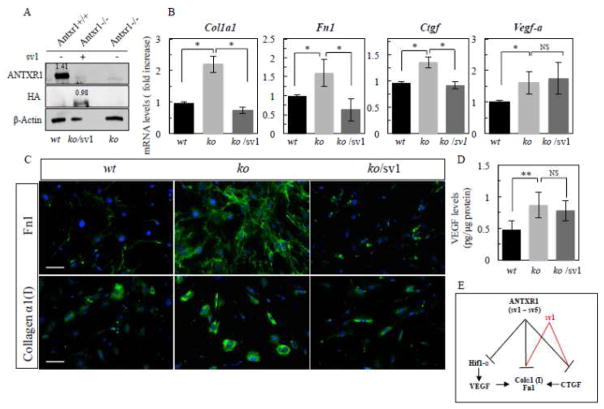
Our previous studies of Antxr1 knockout mice suggested that fibrotic skin abnormalities in these mice are associated with increased VEGF signaling. Here, based on studies of primary fibroblasts isolated from skin of Antx1+/+ and Antxr1-/- mice at embryonic stage E17.5 and postnatal day P49, we conclude that increased Col1a1 and Fn1 expression in Antxr1-deficient fibroblasts is partly mediated by a cell-autonomous ANTXR1-dependent mechanism. In turn, this may act in parallel with VEGF-dependent regulation of collagen type I and fibronectin production. We demonstrate that shRNA mediated knockdown of VEGF in Antxr1-/- fibroblasts reduces Col1a1 and Fn1 expression to below control levels, and these are restored by exogenous addition of recombinant VEGF. In addition, the increase in protein levels of collagen type I and fibronectin in mutant cells is blocked by VEGF neutralizing antibody. However, expressing the longest isoform of ANTXR1 (sv1) in mutant fibroblasts decreases levels of Ctgf, Col1a1 and Fn1 transcripts, but has no effect on VEGF expression. Taken together, our data suggest that the increased matrix production in Antxr1- deficient fibroblasts primarily occurs via a CTGF-dependent pathway and that other ANTXR1-associated mechanisms contribute to VEGF-dependent increase of collagen type I and fibronectin expression. Our findings provide a basis for further studies of novel ANTXR1-dependent connective tissue homeostatic control mechanisms in healthy individuals, patients with organ fibrosis, and patients with GAPO syndrome.
Keywords: ANTXR1/TEM8 GAPO syndrome; CTGF; Collagen type I; Extracellular matrix accumulation; Fibroblasts; Infantile hemangioma; Skin fibrosis; VEGF.
Copyright © 2016 Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Regulatory mechanisms of anthrax toxin receptor 1-dependent vascular and connective tissue homeostasis.Matrix Biol. 2015 Mar;42:56-73. doi: 10.1016/j.matbio.2014.12.002. Epub 2015 Jan 5. Matrix Biol. 2015. PMID: 25572963 Free PMC article.
-
Fell-Muir Lecture: Regulatory mechanisms of skeletal and connective tissue development and homeostasis - lessons from studies of human disorders.Int J Exp Pathol. 2016 Aug;97(4):296-302. doi: 10.1111/iep.12198. Epub 2016 Sep 1. Int J Exp Pathol. 2016. PMID: 27581728 Free PMC article. Review.
-
ALK1 heterozygosity increases extracellular matrix protein expression, proliferation and migration in fibroblasts.Biochim Biophys Acta. 2014 Jun;1843(6):1111-22. doi: 10.1016/j.bbamcr.2014.02.017. Epub 2014 Mar 1. Biochim Biophys Acta. 2014. PMID: 24594380
-
Modulation of collagen type I, fibronectin and dermal fibroblast function and activity, in systemic sclerosis by the antioxidant epigallocatechin-3-gallate.Rheumatology (Oxford). 2010 Nov;49(11):2024-36. doi: 10.1093/rheumatology/keq208. Epub 2010 Jul 13. Rheumatology (Oxford). 2010. PMID: 20627968
-
ANTXR1 as a potential sensor of extracellular mechanical cues.Acta Biomater. 2023 Mar 1;158:80-86. doi: 10.1016/j.actbio.2023.01.006. Epub 2023 Jan 11. Acta Biomater. 2023. PMID: 36638946 Review.
Cited by
-
Mechanism of Action of Monoclonal Antibodies That Block the Activity of the Lethal Toxin of Bacillus Anthracis.Acta Naturae. 2021 Oct-Dec;13(4):98-104. doi: 10.32607/actanaturae.11387. Acta Naturae. 2021. PMID: 35127153 Free PMC article.
-
A large multi-ethnic genome-wide association study identifies novel genetic loci for intraocular pressure.Nat Commun. 2017 Dec 13;8(1):2108. doi: 10.1038/s41467-017-01913-6. Nat Commun. 2017. PMID: 29235454 Free PMC article.
-
CALCR interaction with ANTXR1 drives gastric tumor growth and metastasis via AKT signaling pathway.Sci Rep. 2025 Apr 7;15(1):11826. doi: 10.1038/s41598-025-96310-1. Sci Rep. 2025. PMID: 40195530 Free PMC article.
-
Syndromic and Non-Syndromic Primary Failure of Tooth Eruption: A Genetic Overview.Genes (Basel). 2025 Jan 24;16(2):147. doi: 10.3390/genes16020147. Genes (Basel). 2025. PMID: 40004475 Free PMC article. Review.
-
Latest therapeutic target for gastric cancer: Anthrax toxin receptor 1.World J Gastrointest Oncol. 2021 Apr 15;13(4):216-222. doi: 10.4251/wjgo.v13.i4.216. World J Gastrointest Oncol. 2021. PMID: 33889273 Free PMC article. Review.
References
-
- Werner E, Kowalczyk AP, Faundez V. Anthrax toxin receptor 1/tumor endothelium marker 8 mediates cell spreading by coupling extracellular ligands to the actin cytoskeleton. J Biol Chem. 2006;281:23227–23236. - PubMed
-
- Stranecky V, Hoischen A, Hartmannova H, Zaki MS, Chaudhary A, Zudaire E, Noskova L, Baresova V, Pristoupilova A, Hodanova K, Sovova J, Hulkova H, Piherova L, Hehir-Kwa JY, de Silva D, Senanayake MP, Farrag S, Zeman J, Martasek P, Baxova A, Afifi HH, St Croix B, Brunner HG, Temtamy S, Kmoch S. Mutations in ANTXR1 cause GAPO syndrome. Am J Hum Genet. 2013;92:792–799. - PMC - PubMed
-
- Tipton RE, Gorlin RJ. Growth retardation, alopecia, pseudo-anodontia, and optic atrophy--the GAPO syndrome: report of a patient and review of the literature. Am J Med Genet. 1984;19:209–216. - PubMed
-
- Wajntal A, Koiffmann CP, Mendonca BB, Epps-Quaglia D, Sotto MN, Rati PB, Opitz JM. GAPO syndrome (McKusick 23074)--a connective tissue disorder: report on two affected sibs and on the pathologic findings in the older. Am J Med Genet. 1990;37:213–223. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
