

The Pentatricopeptide Repeat Protein EMB2654 Is Essential for Trans-Splicing of a Chloroplast Small Ribosomal Subunit Transcript
- PMID: 28011633
- PMCID: PMC5291019
- DOI: 10.1104/pp.16.01840
The Pentatricopeptide Repeat Protein EMB2654 Is Essential for Trans-Splicing of a Chloroplast Small Ribosomal Subunit Transcript
Abstract
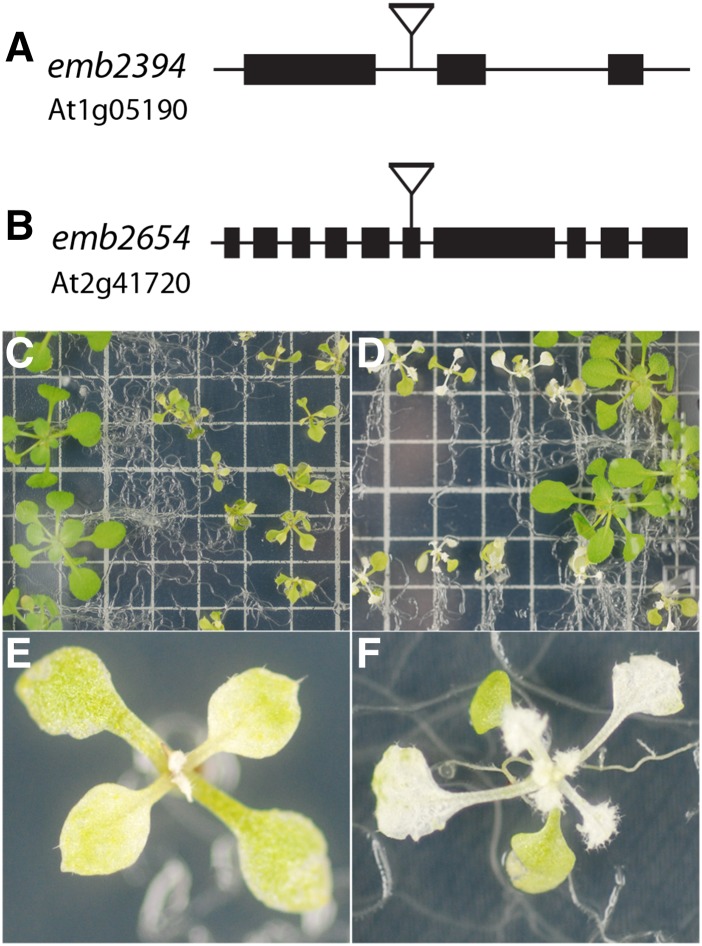
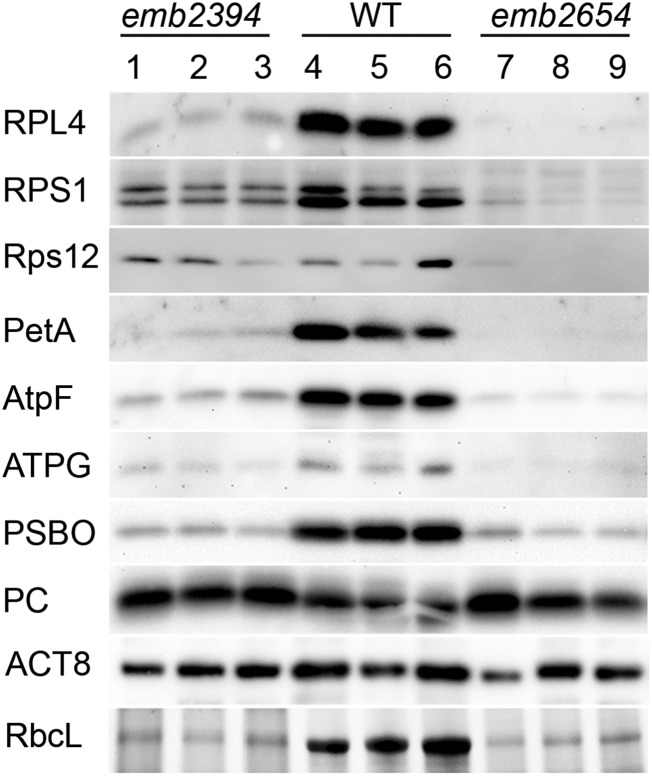
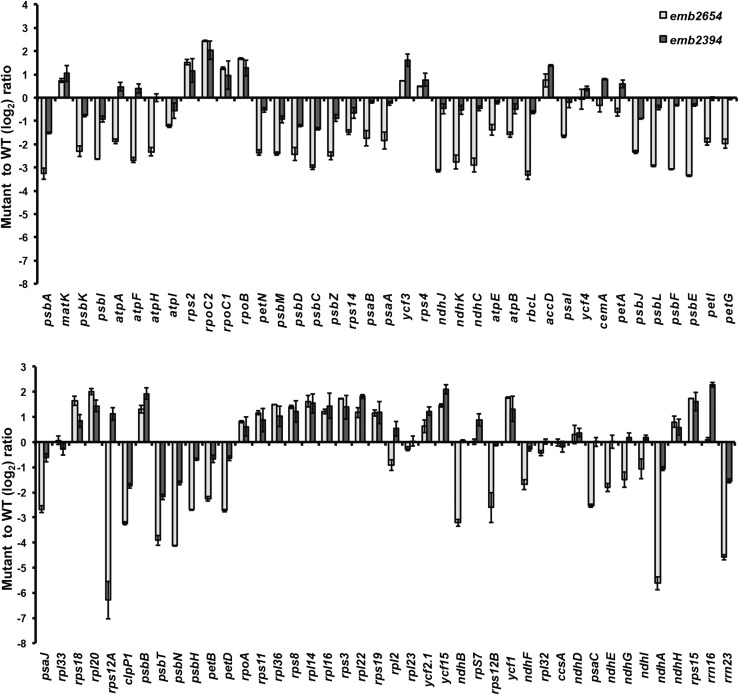
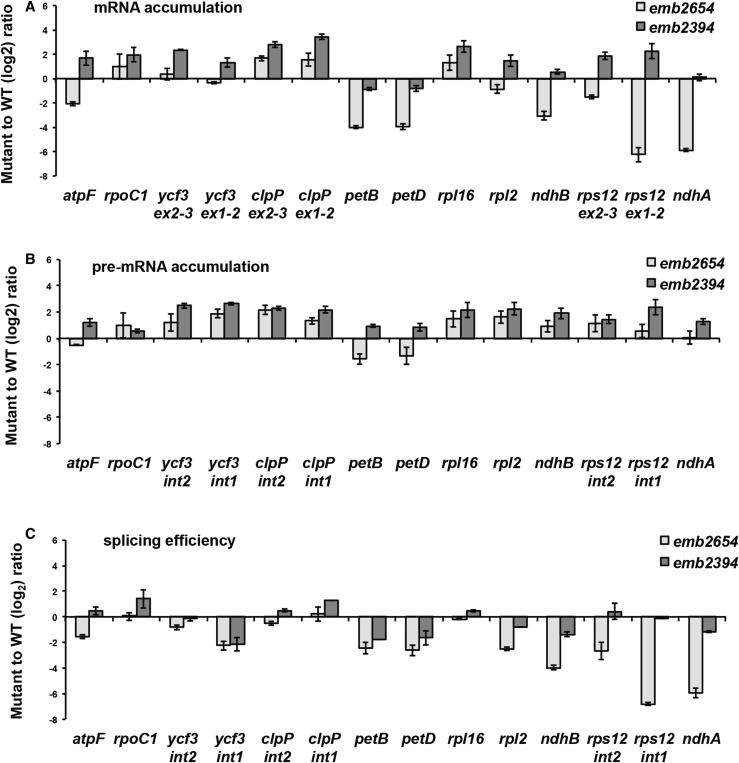
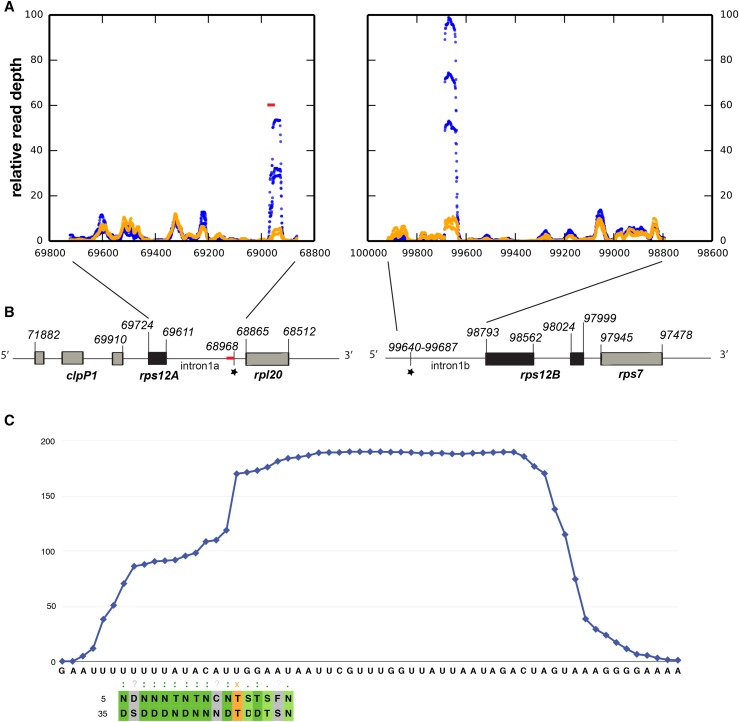
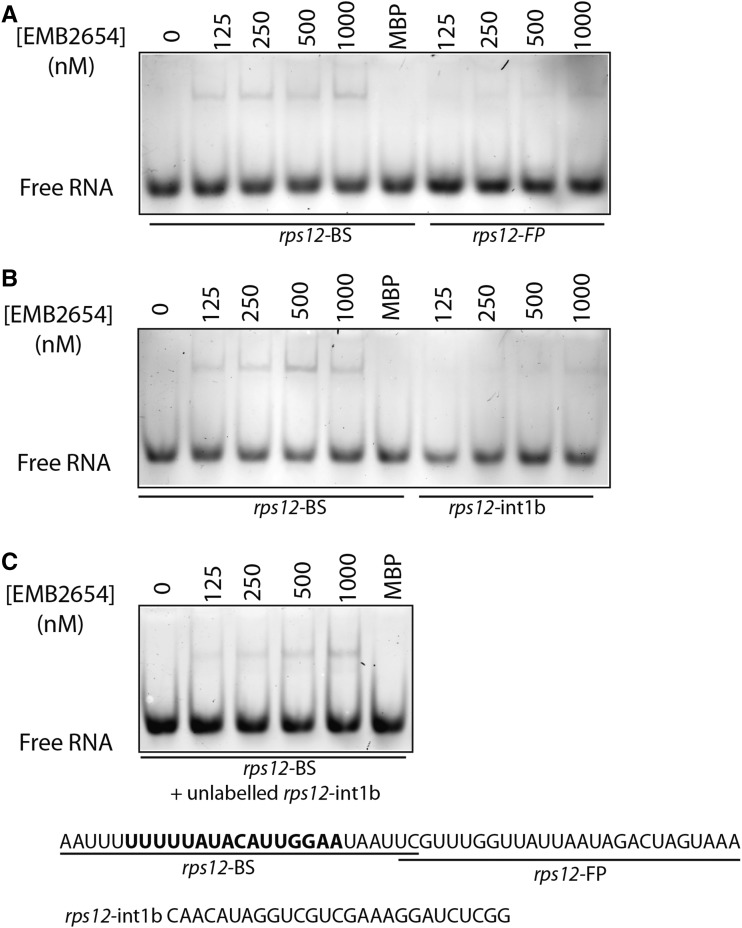
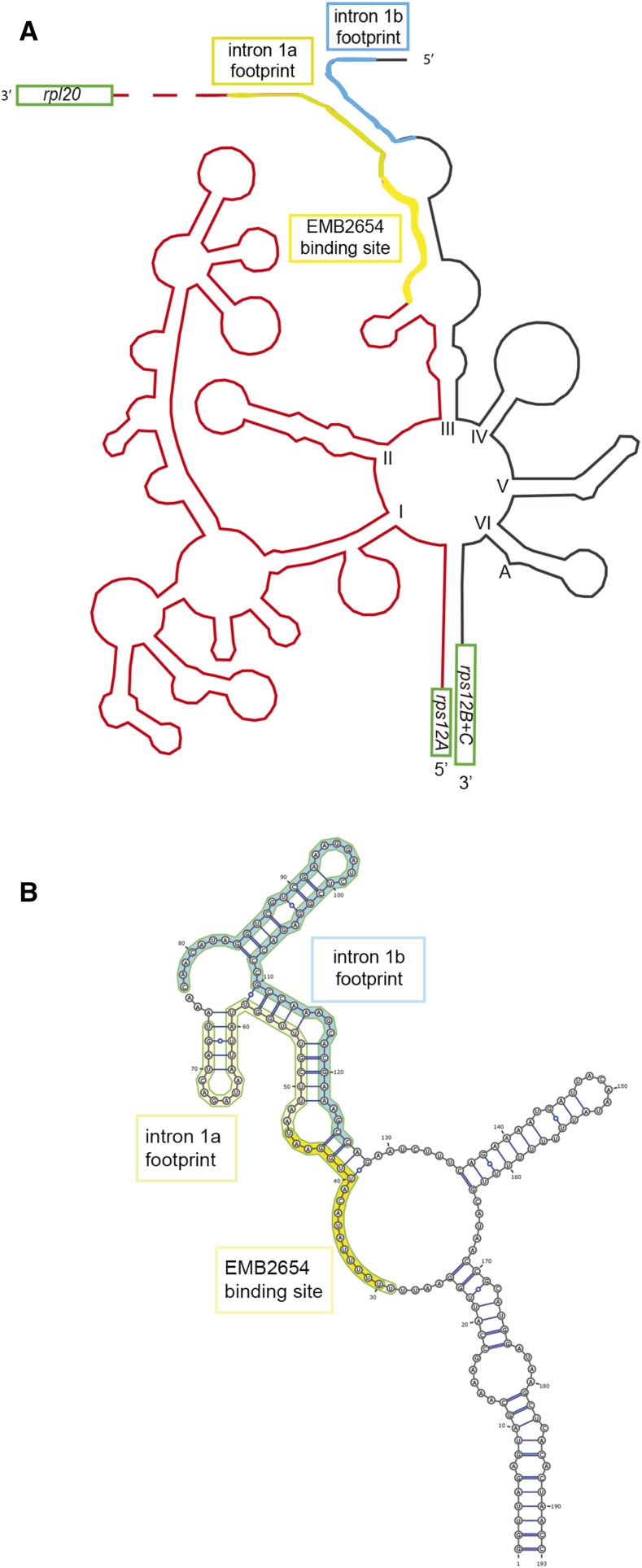
We report the partial complementation and subsequent comparative molecular analysis of two nonviable mutants impaired in chloroplast translation, one (emb2394) lacking the RPL6 protein, and the other (emb2654) carrying a mutation in a gene encoding a P-class pentatricopeptide repeat protein. We show that EMB2654 is required for the trans-splicing of the plastid rps12 transcript and that therefore the emb2654 mutant lacks Rps12 protein and fails to assemble the small subunit of the plastid ribosome, explaining the loss of plastid translation and consequent embryo-lethal phenotype. Predictions of the EMB2654 binding site match a small RNA "footprint" located on the 5' half of the trans-spliced intron that is almost absent in the partially complemented mutant. EMB2654 binds sequence specifically to this target sequence in vitro. Altered patterns in nuclease-protected small RNA fragments in emb2654 show that EMB2654 binding must be an early step in, or prior to, the formation of a large protein-RNA complex covering the free ends of the two rps12 intron halves.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Babiychuk E, Vandepoele K, Wissing J, Garcia-Diaz M, De Rycke R, Akbari H, Joubès J, Beeckman T, Jänsch L, Frentzen M, et al. (2011) Plastid gene expression and plant development require a plastidic protein of the mitochondrial transcription termination factor family. Proc Natl Acad Sci USA 108: 6674–6679 - PMC - PubMed
-
- Barkan A. (2004). Intron splicing in plant organelles. In Daniell H., Chase C.D., eds., Molecular Biology and Biotechnology of Plant Organelles. Springer, Dordrecht, The Netherlands, pp 295–322.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
