

Phosphatidylserine Stimulates Ceramide 1-Phosphate (C1P) Intermembrane Transfer by C1P Transfer Proteins
- PMID: 28011644
- PMCID: PMC5313119
- DOI: 10.1074/jbc.M116.760256
Phosphatidylserine Stimulates Ceramide 1-Phosphate (C1P) Intermembrane Transfer by C1P Transfer Proteins
Abstract
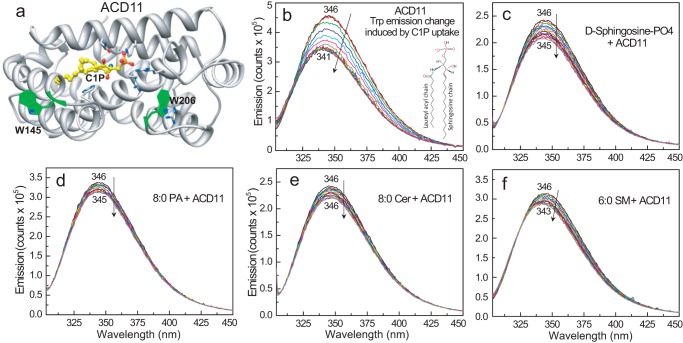
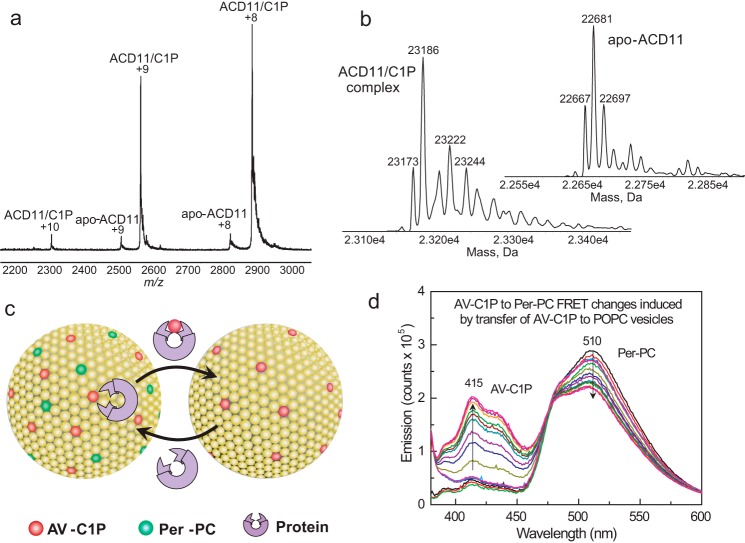
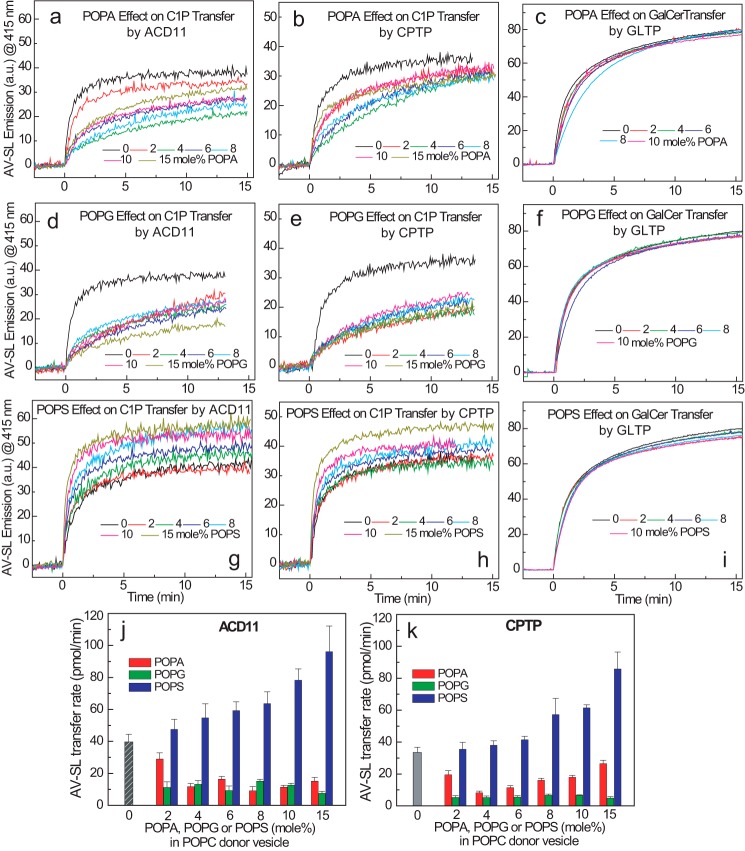
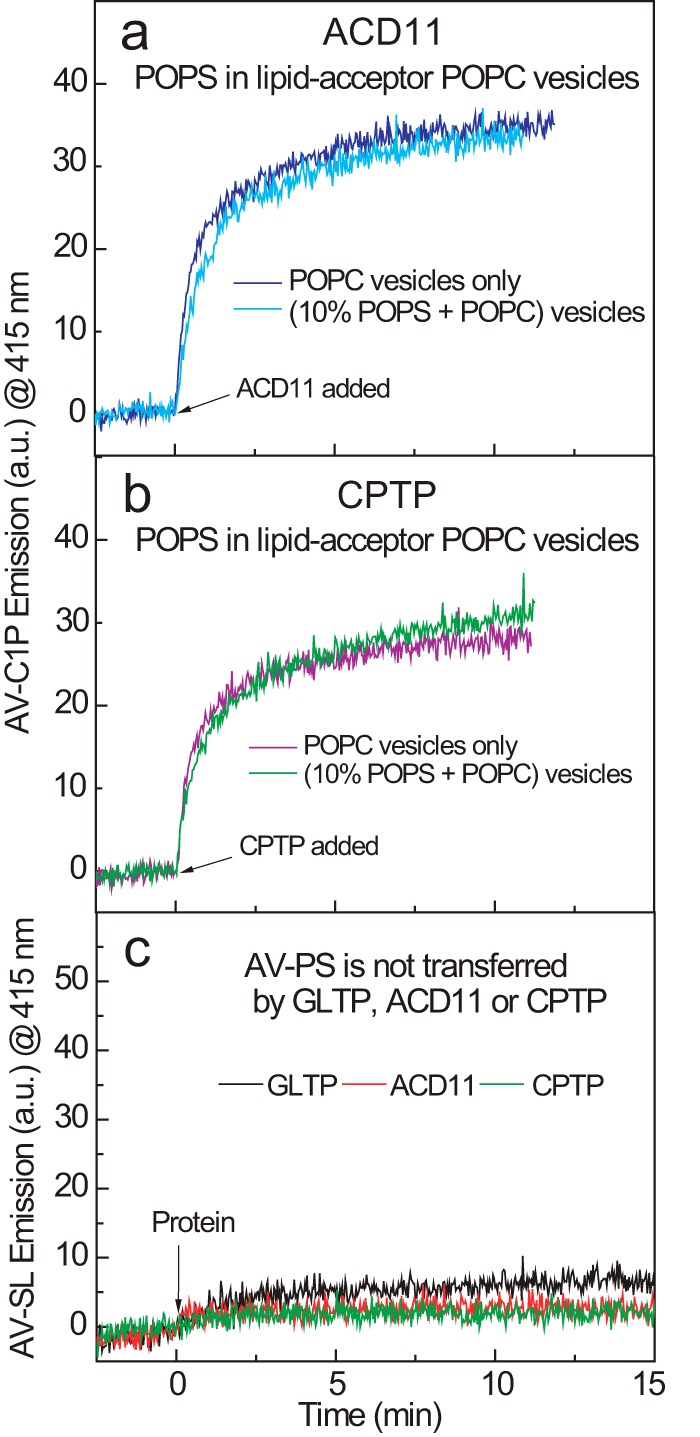
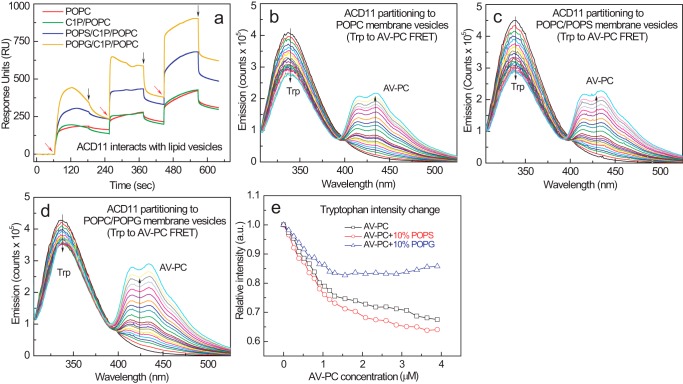
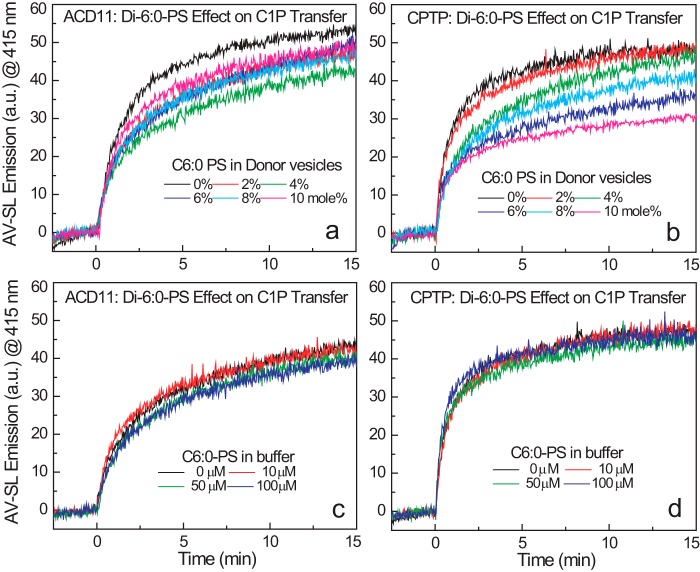
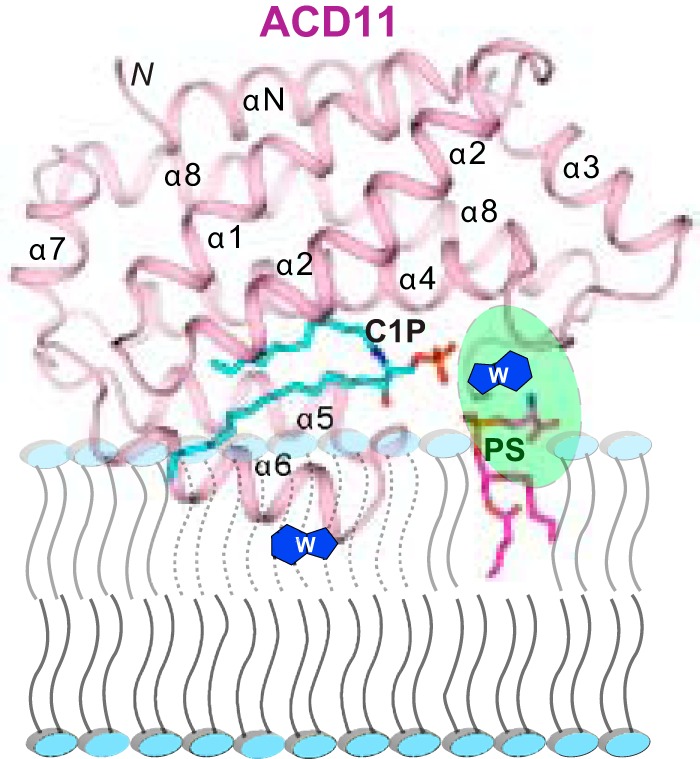
Genetic models for studying localized cell suicide that halt the spread of pathogen infection and immune response activation in plants include Arabidopsis accelerated-cell-death 11 mutant (acd11). In this mutant, sphingolipid homeostasis is disrupted via depletion of ACD11, a lipid transfer protein that is specific for ceramide 1-phosphate (C1P) and phyto-C1P. The C1P binding site in ACD11 and in human ceramide-1-phosphate transfer protein (CPTP) is surrounded by cationic residues. Here, we investigated the functional regulation of ACD11 and CPTP by anionic phosphoglycerides and found that 1-palmitoyl-2-oleoyl-phosphatidic acid or 1-palmitoyl-2-oleoyl-phosphatidylglycerol (≤15 mol %) in C1P source vesicles depressed C1P intermembrane transfer. By contrast, replacement with 1-palmitoyl-2-oleoyl-phosphatidylserine stimulated C1P transfer by ACD11 and CPTP. Notably, "soluble" phosphatidylserine (dihexanoyl-phosphatidylserine) failed to stimulate C1P transfer. Also, none of the anionic phosphoglycerides affected transfer action by human glycolipid lipid transfer protein (GLTP), which is glycolipid-specific and has few cationic residues near its glycolipid binding site. These findings provide the first evidence for a potential phosphoglyceride headgroup-specific regulatory interaction site(s) existing on the surface of any GLTP-fold and delineate new differences between GLTP superfamily members that are specific for C1P versus glycolipid.
Keywords: Arabidopsis thaliana; lipid trafficking; lipid-protein interaction; membrane biophysics; phosphatidic acid; phosphatidylglycerol; phosphatidylserine; protein-lipid interaction; sphingolipid.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Hannun Y. A., and Obeid L. M. (2008) Principles of bioactive lipid signalling: lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 9, 139–150 - PubMed
-
- D'Angelo G., Polishchuk E., Di Tullio G., Santoro M., Di Campli A., Godi A., West G., Bielawski J., Chuang C.-C., van der Spoel A. C., Platt F. M., Hannun Y. A., Polishchuk R., Mattjus P., and De Matteis M. A. (2007) Glycosphingolipid synthesis requires FAPP2 transfer of glucosylceramide. Nature 449, 62–67 - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
