

Protein Phosphatase 2Cs and Microtubule-Associated Stress Protein 1 Control Microtubule Stability, Plant Growth, and Drought Response
- PMID: 28011693
- PMCID: PMC5304354
- DOI: 10.1105/tpc.16.00847
Protein Phosphatase 2Cs and Microtubule-Associated Stress Protein 1 Control Microtubule Stability, Plant Growth, and Drought Response
Abstract
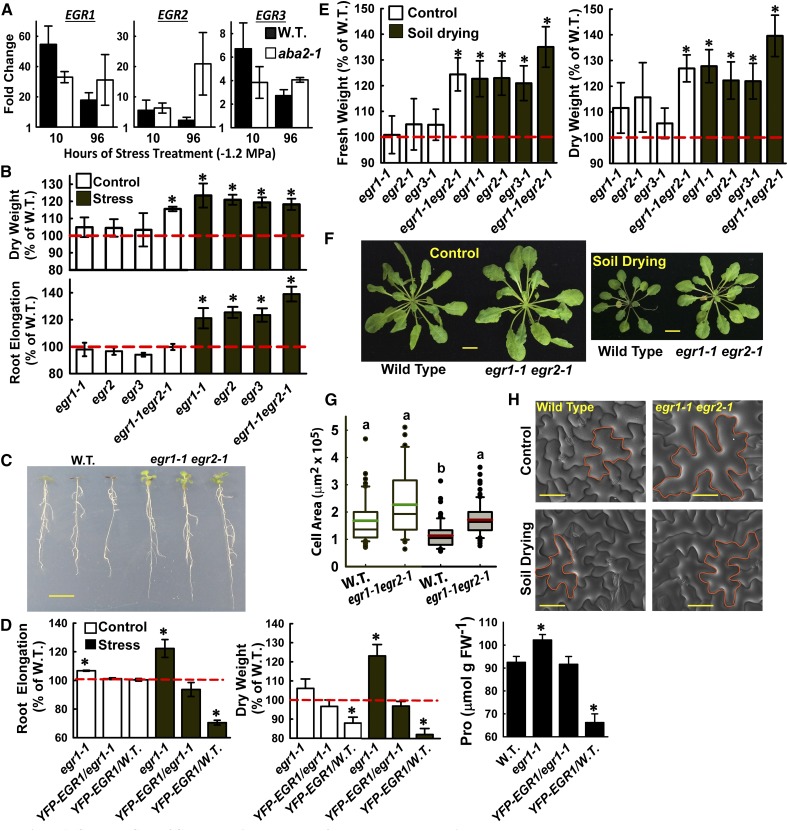
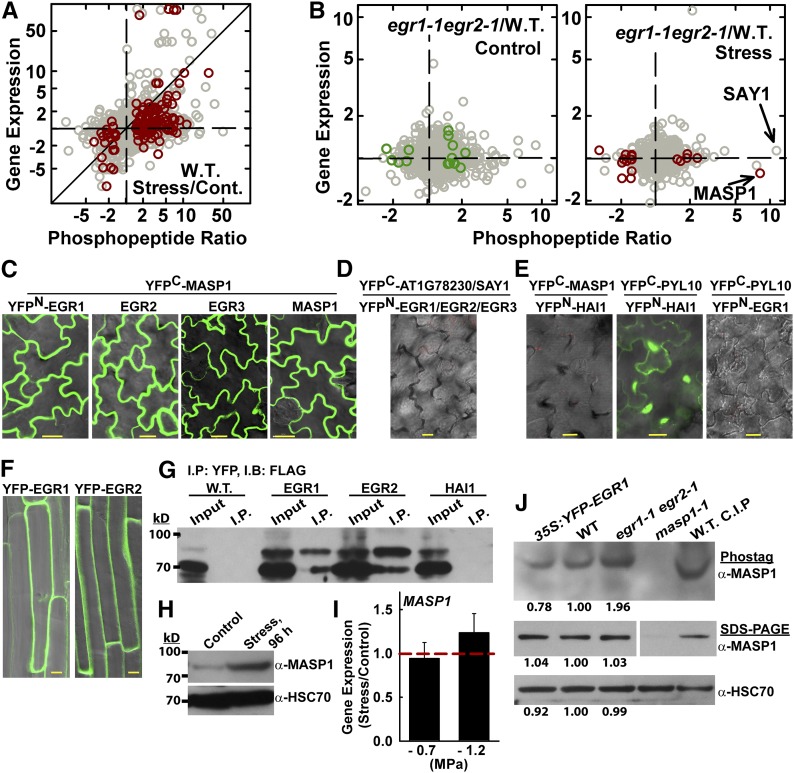
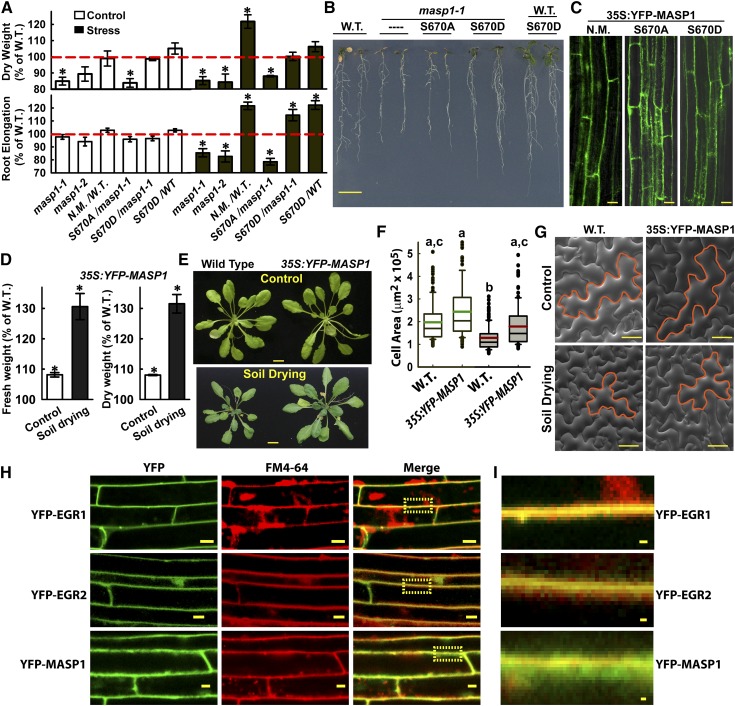
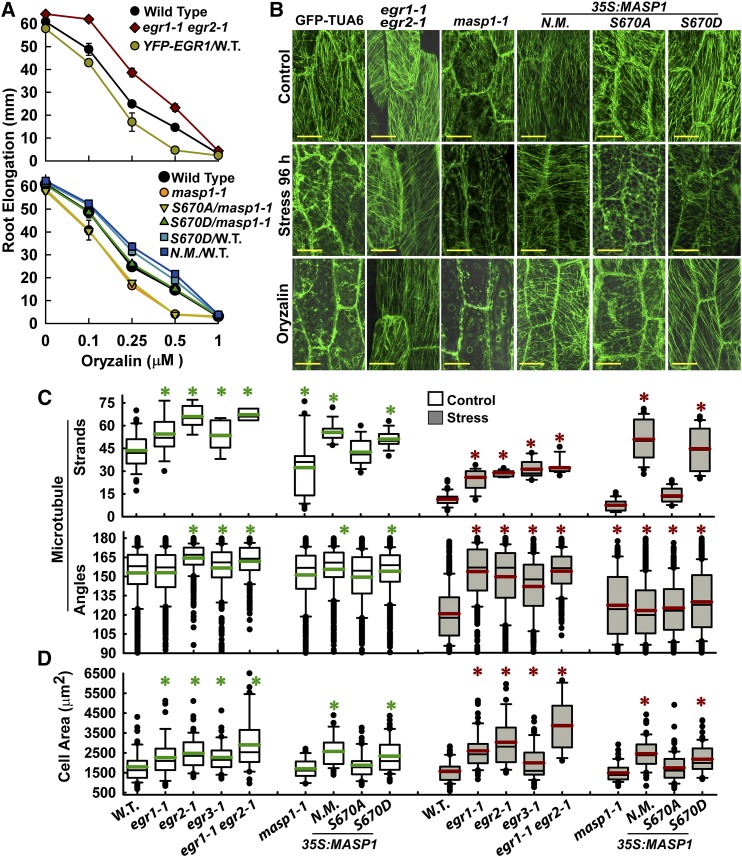
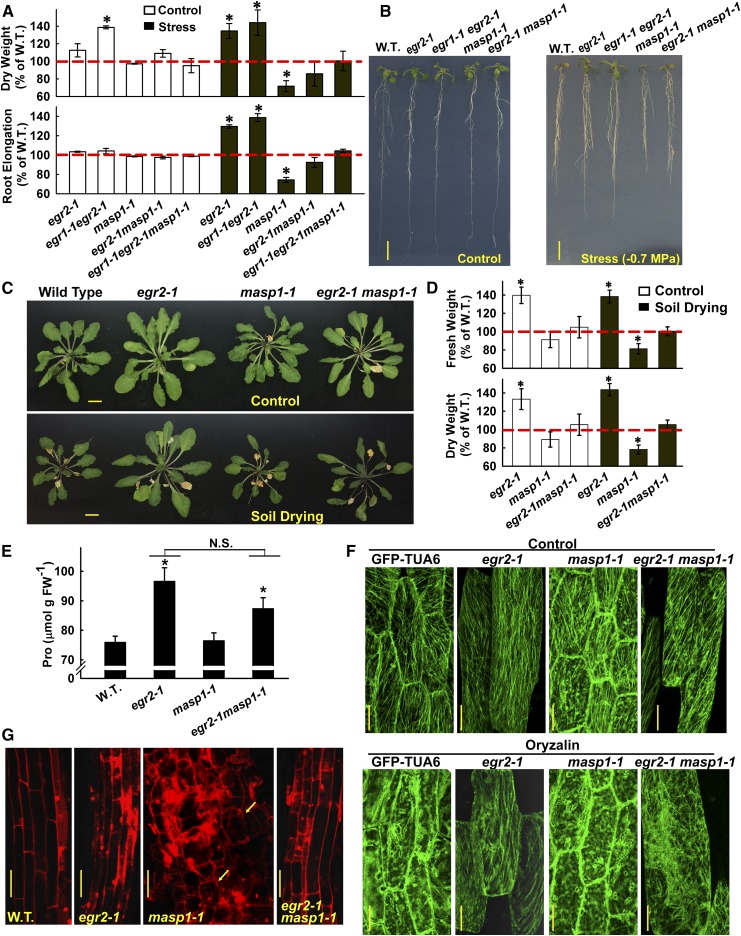
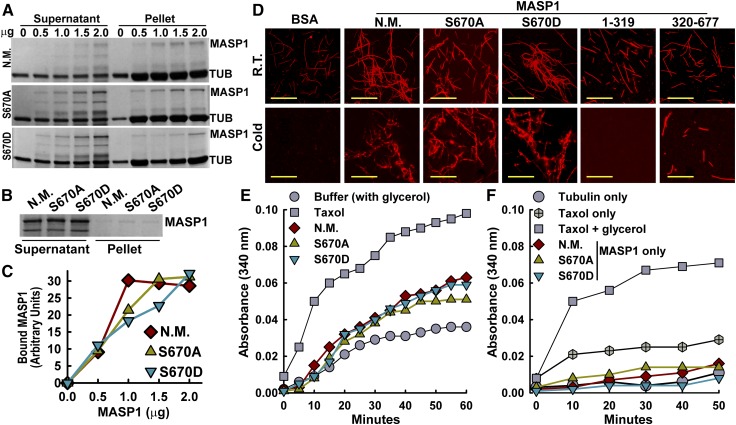
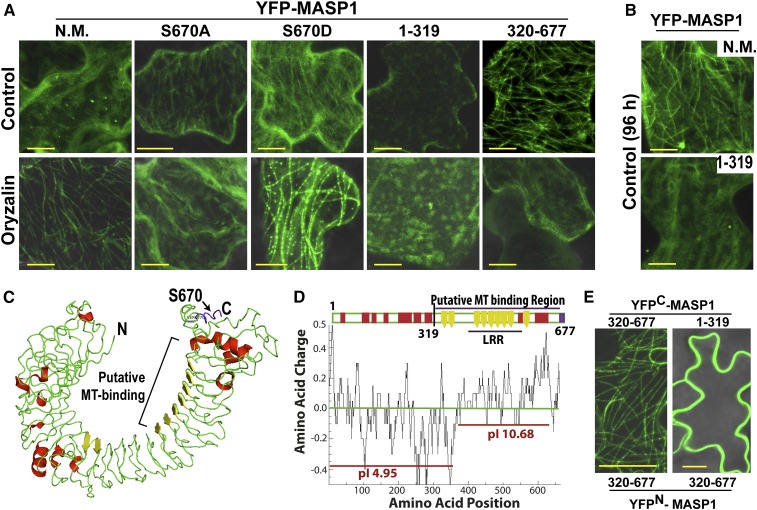
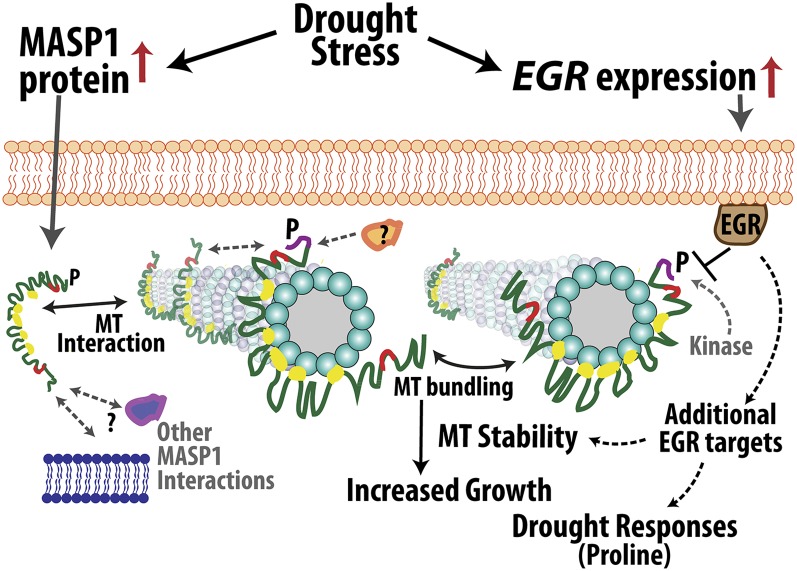
Plant growth is coordinated with environmental factors, including water availability during times of drought. Microtubules influence cell expansion; however, the mechanisms by which environmental signals impinge upon microtubule organization and whether microtubule-related factors limit growth during drought remains unclear. We found that three Clade E Growth-Regulating (EGR) Type 2C protein phosphatases act as negative growth regulators to restrain growth during drought. Quantitative phosphoproteomics indicated that EGRs target cytoskeleton and plasma membrane-associated proteins. Of these, Microtubule-Associated Stress Protein 1 (MASP1), an uncharacterized protein, increased in abundance during stress treatment and could bind, bundle, and stabilize microtubules in vitro. MASP1 overexpression enhanced growth, in vivo microtubule stability, and recovery of microtubule organization during drought acclimation. These MASP1 functions in vivo were dependent on phosphorylation of a single serine. For all EGR and MASP1 mutants and transgenic lines examined, enhanced microtubule recovery and stability were associated with increased growth during drought stress. The EGR-MASP1 system selectively regulates microtubule recovery and stability to adjust plant growth and cell expansion in response to changing environmental conditions. Modification of EGR-MASP1 signaling may be useful to circumvent negative growth regulation limiting plant productivity. EGRs are likely to regulate additional proteins involved in microtubule stability and stress signaling.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures
Comment in
-
Fine-Tuning Plant Growth in the Face of Drought.Plant Cell. 2017 Jan;29(1):4. doi: 10.1105/tpc.17.00038. Epub 2017 Jan 18. Plant Cell. 2017. PMID: 28100705 Free PMC article. No abstract available.
References
-
- Ban Y., Kobayashi Y., Hara T., Hamada T., Hashimoto T., Takeda S., Hattori T. (2013). α-tubulin is rapidly phosphorylated in response to hyperosmotic stress in rice and Arabidopsis. Plant Cell Physiol. 54: 848–858. - PubMed
-
- Bannigan A., Wiedemeier A.M.D., Williamson R.E., Overall R.L., Baskin T.I. (2006). Cortical microtubule arrays lose uniform alignment between cells and are oryzalin resistant in the Arabidopsis mutant, radially swollen 6. Plant Cell Physiol. 47: 949–958. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
