

Coordinated regulation of photosynthetic and respiratory components is necessary to maintain chloroplast energy balance in varied growth conditions
- PMID: 28011719
- PMCID: PMC5441918
- DOI: 10.1093/jxb/erw469
Coordinated regulation of photosynthetic and respiratory components is necessary to maintain chloroplast energy balance in varied growth conditions
Abstract
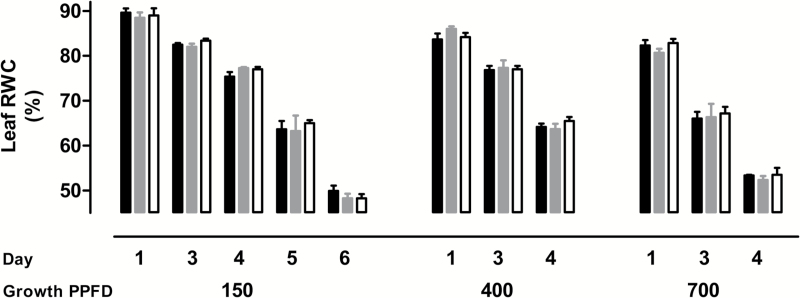
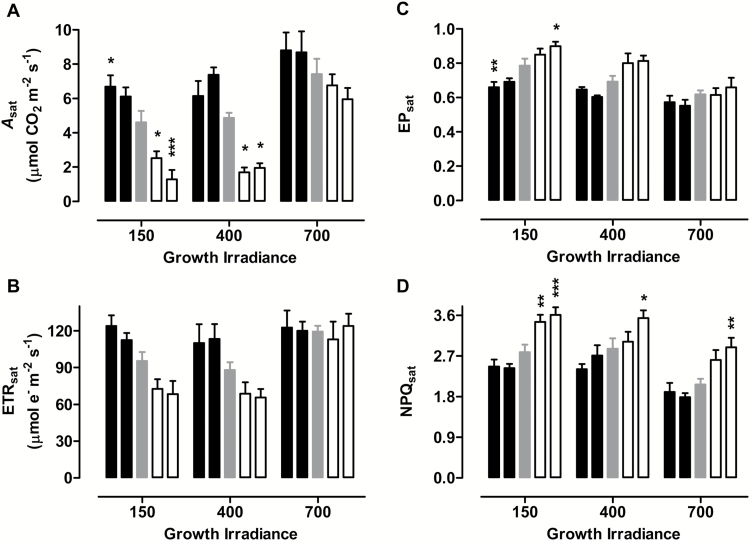
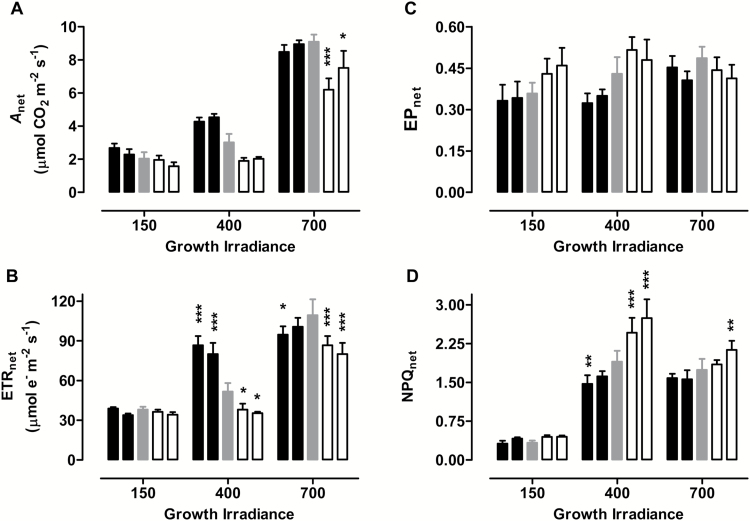
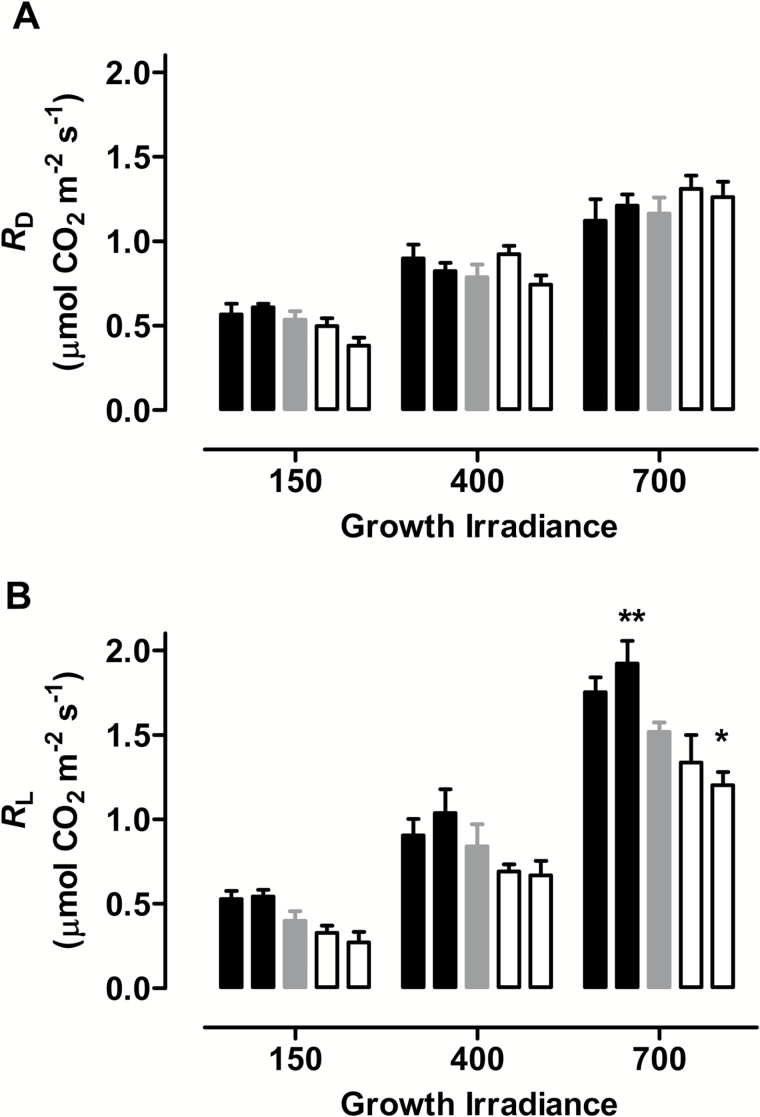
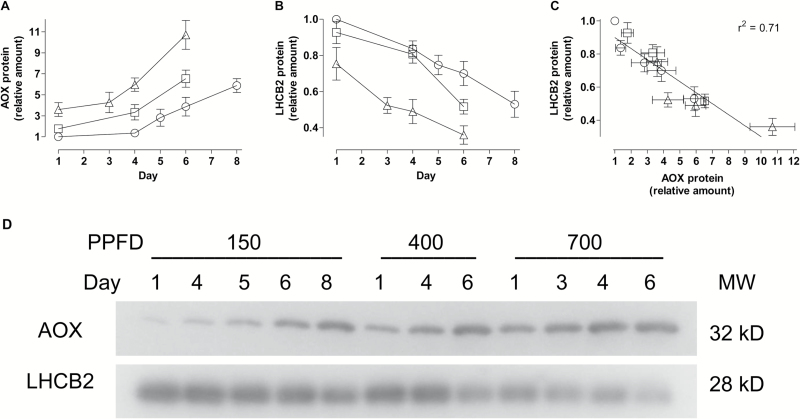
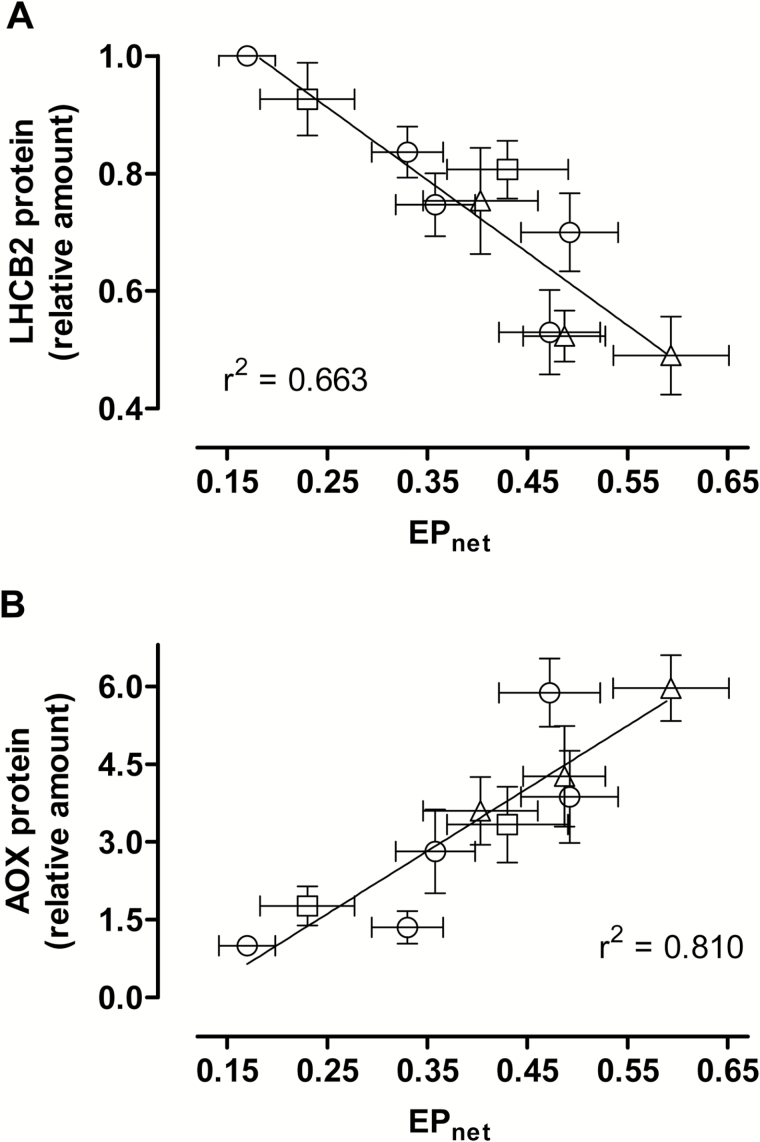
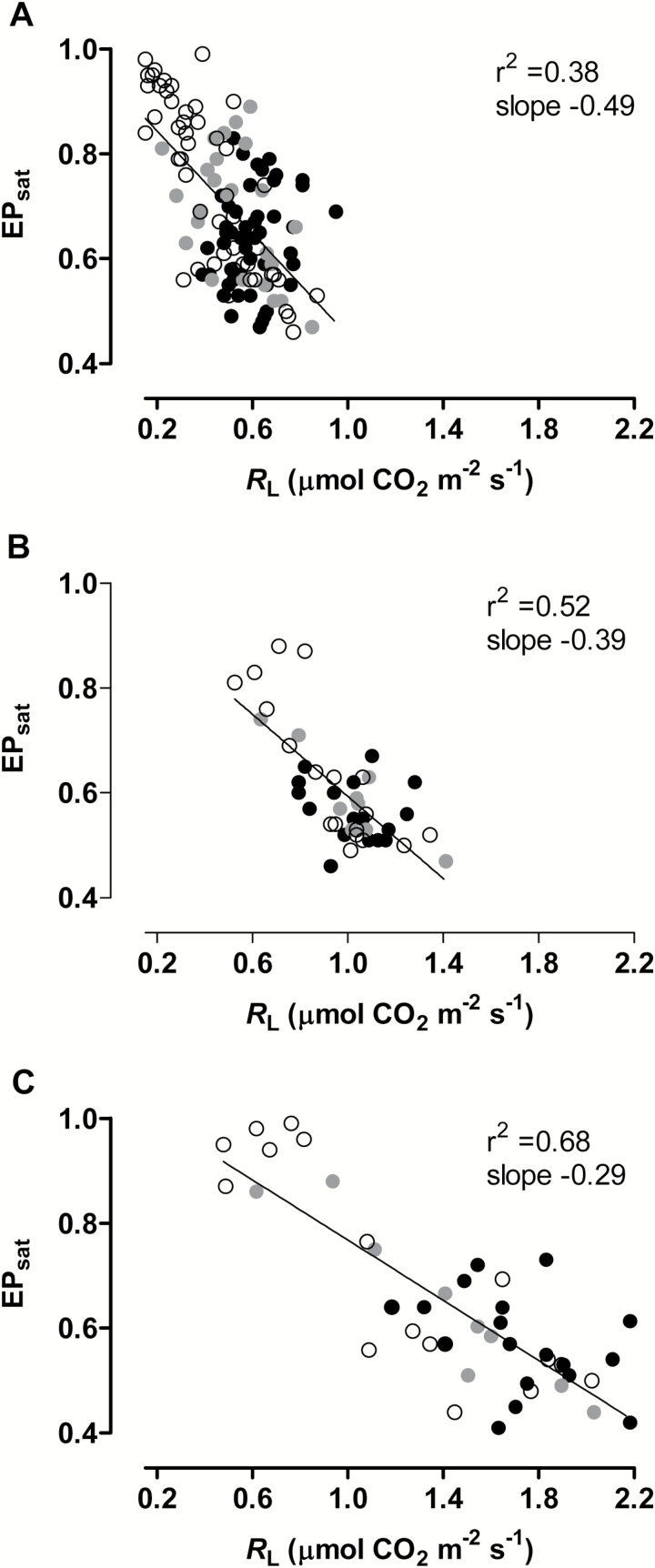
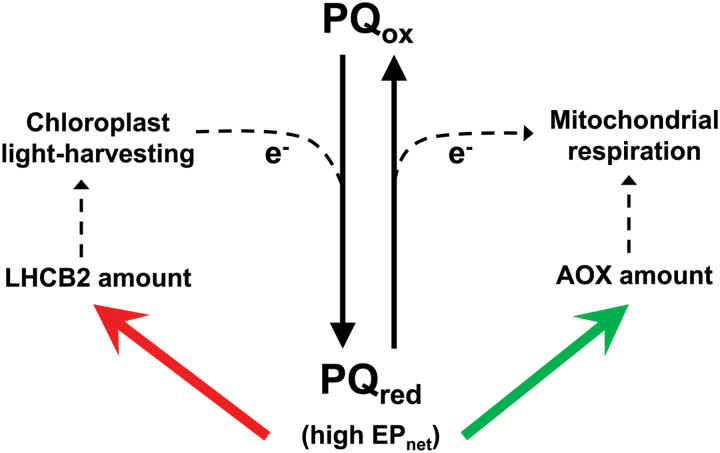
Mitochondria have a non-energy-conserving alternative oxidase (AOX) proposed to support photosynthesis, perhaps by promoting energy balance under varying growth conditions. To investigate this, wild-type (WT) Nicotiana tabacum were compared with AOX knockdown and overexpression lines. In addition, the amount of AOX protein in WT plants was compared with that of chloroplast light-harvesting complex II (LHCB2), whose amount is known to respond to chloroplast energy status. With increased growth irradiance, WT leaves maintained higher rates of respiration in the light (RL), but no differences in RL or photosynthesis were seen between the WT and transgenic lines, suggesting that, under non-stress conditions, AOX was not critical for leaf metabolism, regardless of growth irradiance. However, under drought, the AOX amount became an important determinant of RL, which in turn was an important determinant of chloroplast energy balance (measured as photosystem II excitation pressure, EP), and photosynthetic performance. In the WT, the AOX amount increased and the LHCB2 amount decreased with increased growth irradiance or drought severity. These changes in protein amounts correlated strongly, in opposing ways, with growth EP. This suggests that a signal deriving from the photosynthetic electron transport chain status coordinately controls the amounts of AOX and LHCB2, which then both contribute to maintaining chloroplast energy balance, particularly under stress conditions.
Keywords: Chloroplast energy balance; drought stress; excitation pressure; growth irradiance; light-harvesting complex II protein; mitochondrial alternative oxidase; photosynthesis; respiration.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
Similar articles
-
Alternative oxidase: a respiratory electron transport chain pathway essential for maintaining photosynthetic performance during drought stress.Physiol Plant. 2016 Jul;157(3):322-37. doi: 10.1111/ppl.12451. Epub 2016 May 24. Physiol Plant. 2016. PMID: 27080742 Review.
-
Alternative oxidase respiration maintains both mitochondrial and chloroplast function during drought.New Phytol. 2017 Jan;213(2):560-571. doi: 10.1111/nph.14169. Epub 2016 Aug 31. New Phytol. 2017. PMID: 27579773
-
Improved photosynthetic performance during severe drought in Nicotiana tabacum overexpressing a nonenergy conserving respiratory electron sink.New Phytol. 2015 Oct;208(2):382-95. doi: 10.1111/nph.13479. Epub 2015 May 29. New Phytol. 2015. PMID: 26032897
-
Mitochondrial alternative oxidase maintains respiration and preserves photosynthetic capacity during moderate drought in Nicotiana tabacum.Plant Physiol. 2014 Nov;166(3):1560-74. doi: 10.1104/pp.114.247866. Epub 2014 Sep 9. Plant Physiol. 2014. PMID: 25204647 Free PMC article.
-
Photosynthesis, respiration and growth: A carbon and energy balancing act for alternative oxidase.Mitochondrion. 2020 May;52:197-211. doi: 10.1016/j.mito.2020.04.001. Epub 2020 Apr 9. Mitochondrion. 2020. PMID: 32278748 Review.
Cited by
-
Alternative or cytochrome? Respiratory pathways in traps of aquatic carnivorous bladderwort Utricularia reflexa.Plant Signal Behav. 2022 Dec 31;17(1):2134967. doi: 10.1080/15592324.2022.2134967. Plant Signal Behav. 2022. PMID: 36266991 Free PMC article.
-
Mitochondria in photosynthetic cells: Coordinating redox control and energy balance.Plant Physiol. 2023 Apr 3;191(4):2104-2119. doi: 10.1093/plphys/kiac541. Plant Physiol. 2023. PMID: 36440979 Free PMC article.
-
Co-regulation of mitochondrial and chloroplast function: Molecular components and mechanisms.Plant Commun. 2023 Jan 9;4(1):100496. doi: 10.1016/j.xplc.2022.100496. Epub 2022 Nov 26. Plant Commun. 2023. PMID: 36435968 Free PMC article. Review.
-
Alternative oxidase (AOX) in the carnivorous pitcher plants of the genus Nepenthes: what is it good for?Ann Bot. 2022 Feb 11;129(3):357-365. doi: 10.1093/aob/mcab151. Ann Bot. 2022. PMID: 34922341 Free PMC article.
-
Mitochondrial Uncoupling Protein 1 Overexpression Increases Yield in Nicotiana tabacum under Drought Stress by Improving Source and Sink Metabolism.Front Plant Sci. 2017 Nov 1;8:1836. doi: 10.3389/fpls.2017.01836. eCollection 2017. Front Plant Sci. 2017. PMID: 29163573 Free PMC article.
References
-
- Armstrong AF, Badger MR, Day DA, Barthet MM, Smith PM, Millar AH, Whelan J, Atkin OK. 2008. Dynamic changes in the mitochondrial electron transport chain underpinning cold acclimation of leaf respiration. Plant, Cell & Environment 31, 1156–1169. - PubMed
-
- Ayub G, Smith RA, Tissue DT, Atkin OK. 2011. Impacts of drought on leaf respiration in darkness and light in Eucalyptus saligna exposed to industrial-age atmospheric CO2 and growth temperature. The New Phytologist 190, 1003–1018. - PubMed
-
- Bailey S, Walters RG, Jansson S, Horton P. 2001. Acclimation of Arabidopsis thaliana to the light environment: the existence of separate low light and high light responses. Planta 213, 794–801. - PubMed
-
- Bartoli CG, Yu J, Gómez F, Fernández L, McIntosh L, Foyer CH. 2006. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. Journal of Experimental Botany 57, 1621–1631. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
