

Differential distribution and functional impact of BK channel beta1 subunits across mesenteric, coronary, and different cerebral arteries of the rat
- PMID: 28012000
- PMCID: PMC5432129
- DOI: 10.1007/s00424-016-1929-z
Differential distribution and functional impact of BK channel beta1 subunits across mesenteric, coronary, and different cerebral arteries of the rat
Abstract
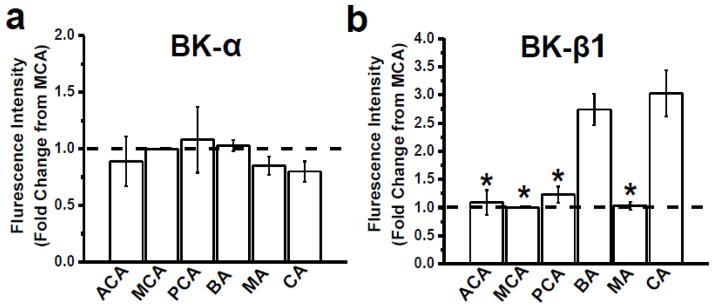
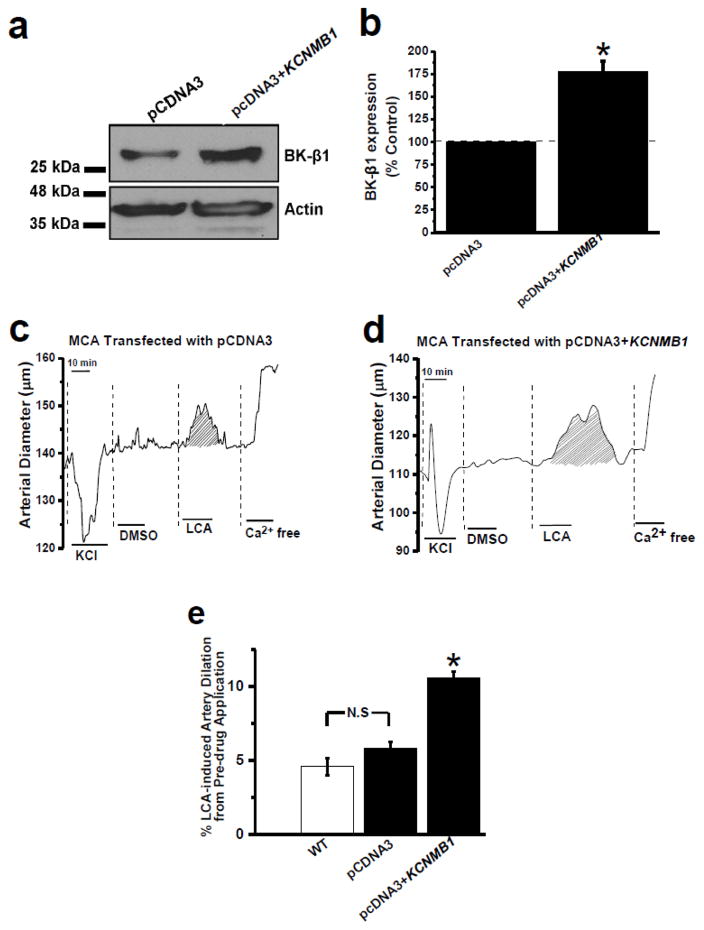
Large conductance, Ca2+i- and voltage-gated K+ (BK) channels regulate myogenic tone and, thus, arterial diameter. In smooth muscle (SM), BK channels include channel-forming α and auxiliary β1 subunits. BK β1 increases the channel's Ca2+ sensitivity, allowing BK channels to negatively feedback on depolarization-induced Ca2+ entry, oppose SM contraction and favor vasodilation. Thus, endothelial-independent vasodilation can be evoked though targeting of SM BK β1 by endogenous ligands, including lithocholate (LCA). Here, we investigated the expression of BK β1 across arteries of the cerebral and peripheral circulations, and the contribution of such expression to channel function and BK β1-mediated vasodilation. Data demonstrate that endothelium-independent, BK β1-mediated vasodilation by LCA is larger in coronary (CA) and basilar (BA) arteries than in anterior cerebral (ACA), middle cerebral (MCA), posterior cerebral (PCA), and mesenteric (MA) arteries, all arterial segments having a similar diameter. Thus, differential dilation occurs in extracranial arteries which are subjected to similar vascular pressure (CA vs. MA) and in arteries that irrigate different brain regions (BA vs. ACA, MCA, and PCA). SM BK channels from BA and CA displayed increased basal activity and LCA responses, indicating increased BK β1 functional presence. Indeed, in the absence of detectable changes in BK α, BA and CA myocytes showed an increased location of BK β1 in the plasmalemma/subplasmalemma. Moreover, these myocytes distinctly showed increased BK β1 messenger RNA (mRNA) levels. Supporting a major role of enhanced BK β1 transcripts in artery dilation, LCA-induced dilation of MCA transfected with BK β1 complementary DNA (cDNA) was as high as LCA-induced dilation of untransfected BA or CA.
Keywords: BK β1 subunit; Cerebral artery myocyte; KCNMB1; Lithocholate; MaxiK channel; Vascular smooth muscle; Vasodilation.
Figures






Similar articles
-
Cholesterol activates BK channels by increasing KCNMB1 protein levels in the plasmalemma.J Biol Chem. 2021 Jan-Jun;296:100381. doi: 10.1016/j.jbc.2021.100381. Epub 2021 Feb 6. J Biol Chem. 2021. PMID: 33556372 Free PMC article.
-
Cerebrovascular dilation via selective targeting of the cholane steroid-recognition site in the BK channel β1-subunit by a novel nonsteroidal agent.Mol Pharmacol. 2013 May;83(5):1030-44. doi: 10.1124/mol.112.083519. Epub 2013 Mar 1. Mol Pharmacol. 2013. PMID: 23455312 Free PMC article.
-
Equol increases cerebral blood flow in rats via activation of large-conductance Ca(2+)-activated K(+) channels in vascular smooth muscle cells.Pharmacol Res. 2016 May;107:186-194. doi: 10.1016/j.phrs.2016.03.015. Epub 2016 Mar 16. Pharmacol Res. 2016. PMID: 26995303
-
Reduction of large-conductance Ca²(+) -activated K(+) channel with compensatory increase of nitric oxide in insulin resistant rats.Diabetes Metab Res Rev. 2011 Jul;27(5):461-9. doi: 10.1002/dmrr.1196. Diabetes Metab Res Rev. 2011. PMID: 21425425 Review.
-
Functional regulation of large conductance Ca2+-activated K+ channels in vascular diseases.Metabolism. 2018 Jun;83:75-80. doi: 10.1016/j.metabol.2018.01.008. Epub 2018 Jan 31. Metabolism. 2018. PMID: 29373813 Review.
Cited by
-
Regulation of BK Channels by Beta and Gamma Subunits.Annu Rev Physiol. 2019 Feb 10;81:113-137. doi: 10.1146/annurev-physiol-022516-034038. Annu Rev Physiol. 2019. PMID: 30742788 Free PMC article. Review.
-
Cholesterol activates BK channels by increasing KCNMB1 protein levels in the plasmalemma.J Biol Chem. 2021 Jan-Jun;296:100381. doi: 10.1016/j.jbc.2021.100381. Epub 2021 Feb 6. J Biol Chem. 2021. PMID: 33556372 Free PMC article.
-
Molecular structures of the human Slo1 K+ channel in complex with β4.Elife. 2019 Dec 9;8:e51409. doi: 10.7554/eLife.51409. Elife. 2019. PMID: 31815672 Free PMC article.
-
Toluene is a cerebral artery constrictor acting via BK channels.Neuropharmacology. 2025 Mar 15;266:110272. doi: 10.1016/j.neuropharm.2024.110272. Epub 2024 Dec 18. Neuropharmacology. 2025. PMID: 39706291
-
Progesterone activation of β1-containing BK channels involves two binding sites.Nat Commun. 2023 Nov 9;14(1):7248. doi: 10.1038/s41467-023-42827-w. Nat Commun. 2023. PMID: 37945687 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
