

Interaction of E. coli Hsp90 with DnaK Involves the DnaJ Binding Region of DnaK
- PMID: 28013030
- PMCID: PMC5357148
- DOI: 10.1016/j.jmb.2016.12.014
Interaction of E. coli Hsp90 with DnaK Involves the DnaJ Binding Region of DnaK
Abstract
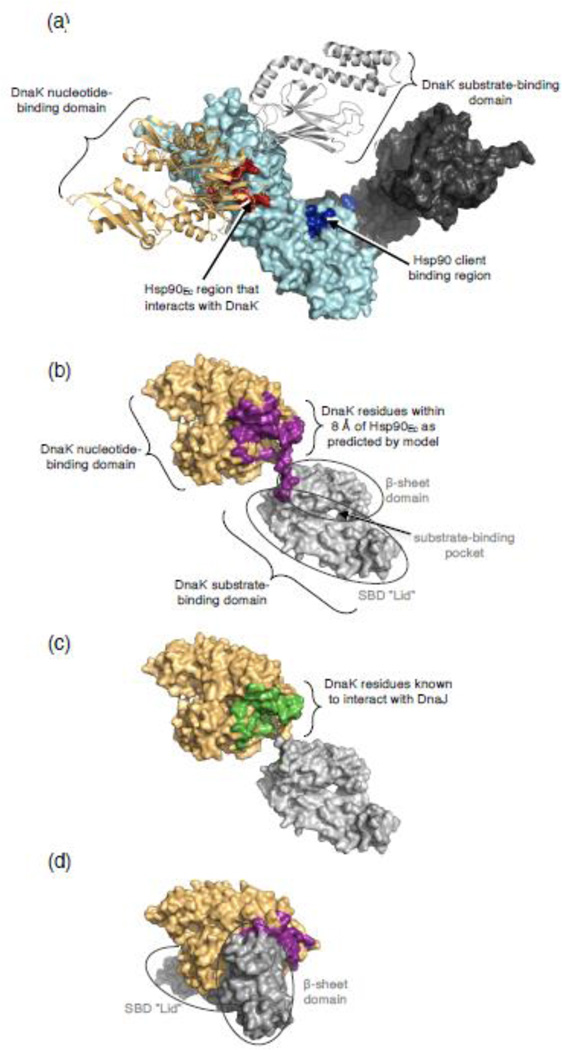
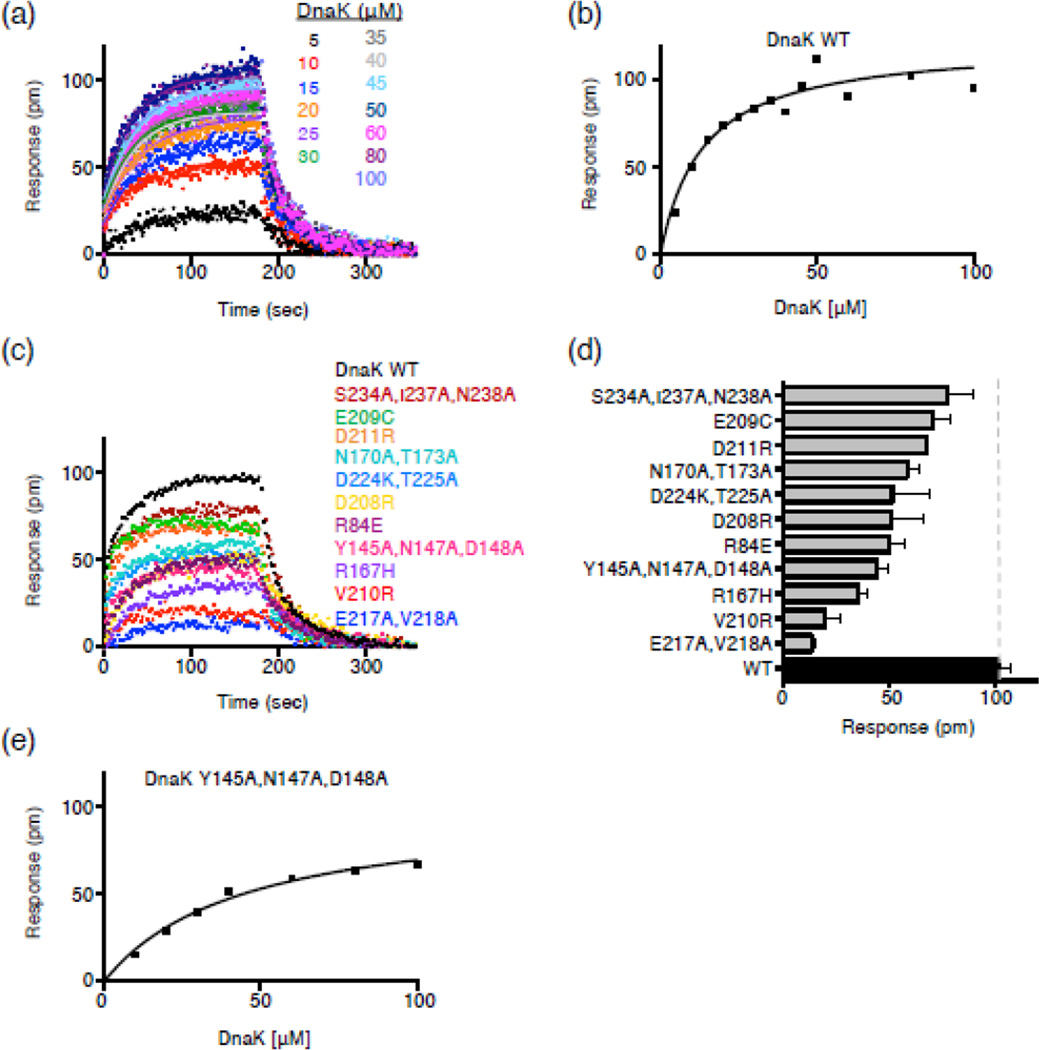
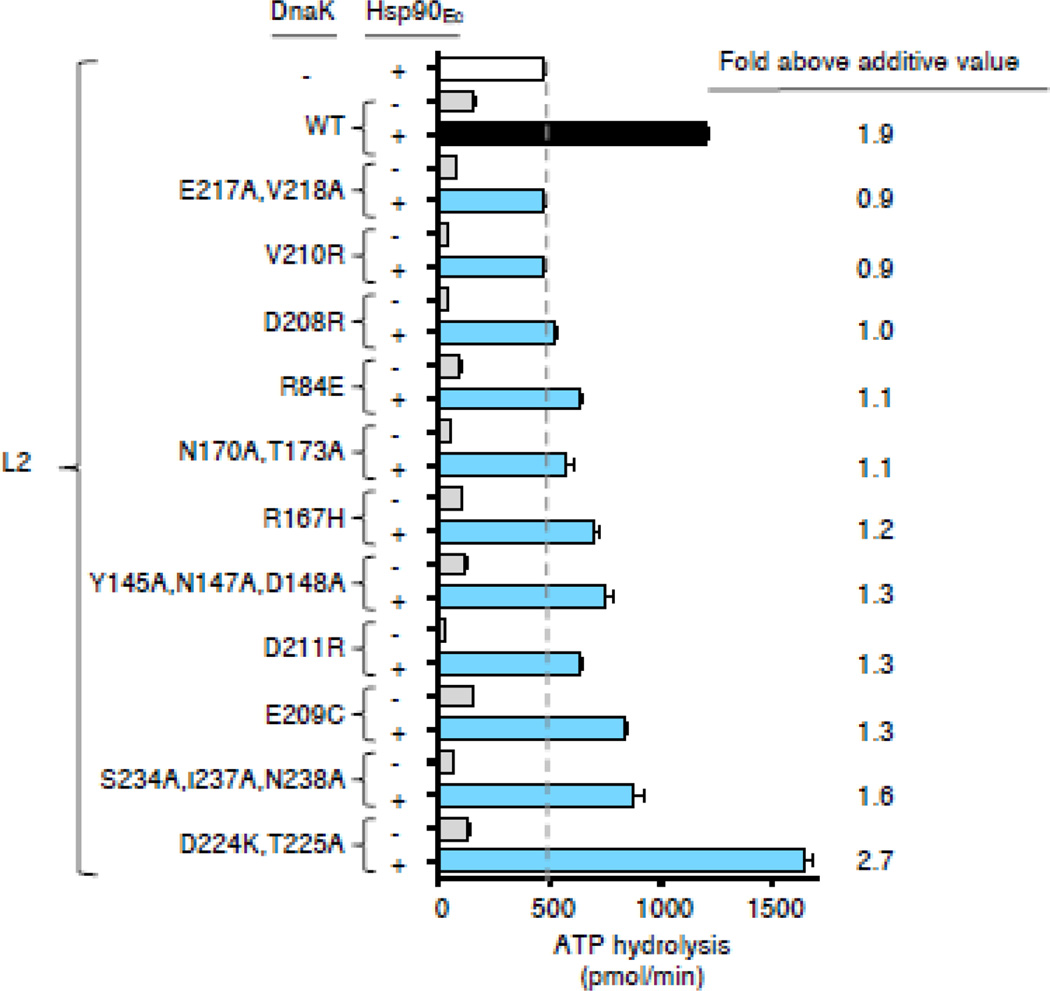
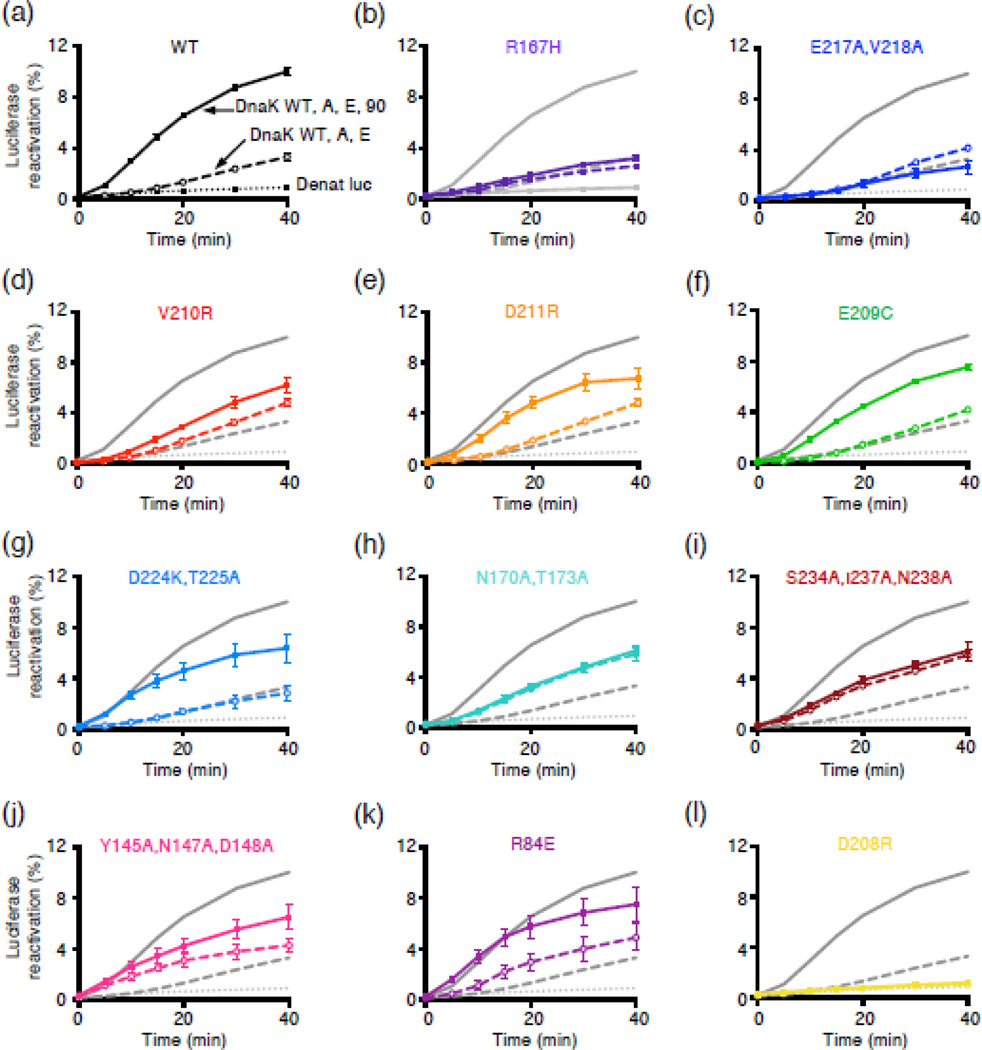
The 90-kDa heat shock protein (Hsp90) is a widely conserved and ubiquitous molecular chaperone that participates in ATP-dependent protein remodeling in both eukaryotes and prokaryotes. It functions in conjunction with Hsp70 and the Hsp70 cochaperones, an Hsp40 (J-protein) and a nucleotide exchange factor. In Escherichia coli, the functional collaboration between Hsp90Ec and Hsp70, DnaK, requires that the two chaperones directly interact. We used molecular docking to model the interaction of Hsp90Ec and DnaK. The top-ranked docked model predicted that a region in the nucleotide-binding domain (NBD) of DnaK interacted with a region in the middle domain of Hsp90Ec. We then made substitution mutants in DnaK residues suggested by the model to interact with Hsp90Ec. Of the 12 mutants tested, 11 were defective or partially defective in their ability to interact with Hsp90Ecin vivo in a bacterial two-hybrid assay and in vitro in a bio-layer interferometry assay. These DnaK mutants were also defective in their ability to function collaboratively in protein remodeling with Hsp90Ec but retained the ability to act with DnaK cochaperones. Taken together, these results suggest that a specific region in the NBD of DnaK is involved in the interaction with Hsp90Ec, and this interaction is functionally important. Moreover, the region of DnaK that we found to be necessary for Hsp90Ec binding includes residues that are also involved in J-protein binding, suggesting a functional interplay among DnaK, DnaK cochaperones, and Hsp90Ec.
Keywords: CbpA; Hsp40; HtpG; molecular chaperone; protein remodeling.
Published by Elsevier Ltd.
Figures


Similar articles
-
Hsp70 and Hsp90 of E. coli Directly Interact for Collaboration in Protein Remodeling.J Mol Biol. 2015 Dec 4;427(24):3877-89. doi: 10.1016/j.jmb.2015.10.010. Epub 2015 Oct 23. J Mol Biol. 2015. PMID: 26482100 Free PMC article.
-
Structural Communication between the E. coli Chaperones DnaK and Hsp90.Int J Mol Sci. 2021 Feb 23;22(4):2200. doi: 10.3390/ijms22042200. Int J Mol Sci. 2021. PMID: 33672263 Free PMC article.
-
Hsp90, DnaK, and ClpB collaborate in protein reactivation.Proc Natl Acad Sci U S A. 2025 Feb 4;122(5):e2422640122. doi: 10.1073/pnas.2422640122. Epub 2025 Jan 29. Proc Natl Acad Sci U S A. 2025. PMID: 39879241 Free PMC article.
-
Hsp90 and Hsp70 chaperones: Collaborators in protein remodeling.J Biol Chem. 2019 Feb 8;294(6):2109-2120. doi: 10.1074/jbc.REV118.002806. Epub 2018 Nov 6. J Biol Chem. 2019. PMID: 30401745 Free PMC article. Review.
-
The Bacterial Hsp90 Chaperone: Cellular Functions and Mechanism of Action.Annu Rev Microbiol. 2021 Oct 8;75:719-739. doi: 10.1146/annurev-micro-032421-035644. Epub 2021 Aug 10. Annu Rev Microbiol. 2021. PMID: 34375543 Review.
Cited by
-
Bacterial Hsp90 Facilitates the Degradation of Aggregation-Prone Hsp70-Hsp40 Substrates.Front Mol Biosci. 2021 Apr 15;8:653073. doi: 10.3389/fmolb.2021.653073. eCollection 2021. Front Mol Biosci. 2021. PMID: 33937334 Free PMC article.
-
The Yeast Hsp70 Cochaperone Ydj1 Regulates Functional Distinction of Ssa Hsp70s in the Hsp90 Chaperoning Pathway.Genetics. 2020 Jul;215(3):683-698. doi: 10.1534/genetics.120.303190. Epub 2020 Apr 16. Genetics. 2020. PMID: 32299842 Free PMC article.
-
Modeling Hsp70/Hsp40 interaction by multi-scale molecular simulations and coevolutionary sequence analysis.Elife. 2017 May 12;6:e23471. doi: 10.7554/eLife.23471. Elife. 2017. PMID: 28498104 Free PMC article.
-
The Hsp70-Hsp90 co-chaperone Hop/Stip1 shifts the proteostatic balance from folding towards degradation.Nat Commun. 2020 Nov 25;11(1):5975. doi: 10.1038/s41467-020-19783-w. Nat Commun. 2020. PMID: 33239621 Free PMC article.
-
Ydj1 interaction at nucleotide-binding-domain of yeast Ssa1 impacts Hsp90 collaboration and client maturation.PLoS Genet. 2022 Nov 9;18(11):e1010442. doi: 10.1371/journal.pgen.1010442. eCollection 2022 Nov. PLoS Genet. 2022. PMID: 36350833 Free PMC article.
References
-
- Johnson JL. Evolution and function of diverse Hsp90 homologs and cochaperone proteins. Biochim. Biophys. Acta. - Molecular Cell Research. 2012;1823:607–613. - PubMed
-
- Mayer MP. Gymnastics of molecular chaperones. Mol. Cell. 2010;39:321–331. - PubMed
-
- Röhl A, Rohrberg J, Buchner J. The chaperone Hsp90: Changing partners for demanding clients. Trends Biochem. Sci. 2013;38:253–262. - PubMed
-
- Taipale M, Jarosz DF, Lindquist S. HSP90 at the hub of protein homeostasis: emerging mechanistic insights. Nat. Rev. Mol. Cell Biol. 2010;11:515–528. - PubMed
-
- Li J, Soroka J, Buchner J. The Hsp90 chaperone machinery: Conformational dynamics and regulation by co-chaperones. Biochim. Biophys. Acta. 2012;1823:624–635. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
