

An Actin Network Dispatches Ciliary GPCRs into Extracellular Vesicles to Modulate Signaling
- PMID: 28017328
- PMCID: PMC5235987
- DOI: 10.1016/j.cell.2016.11.036
An Actin Network Dispatches Ciliary GPCRs into Extracellular Vesicles to Modulate Signaling
Abstract
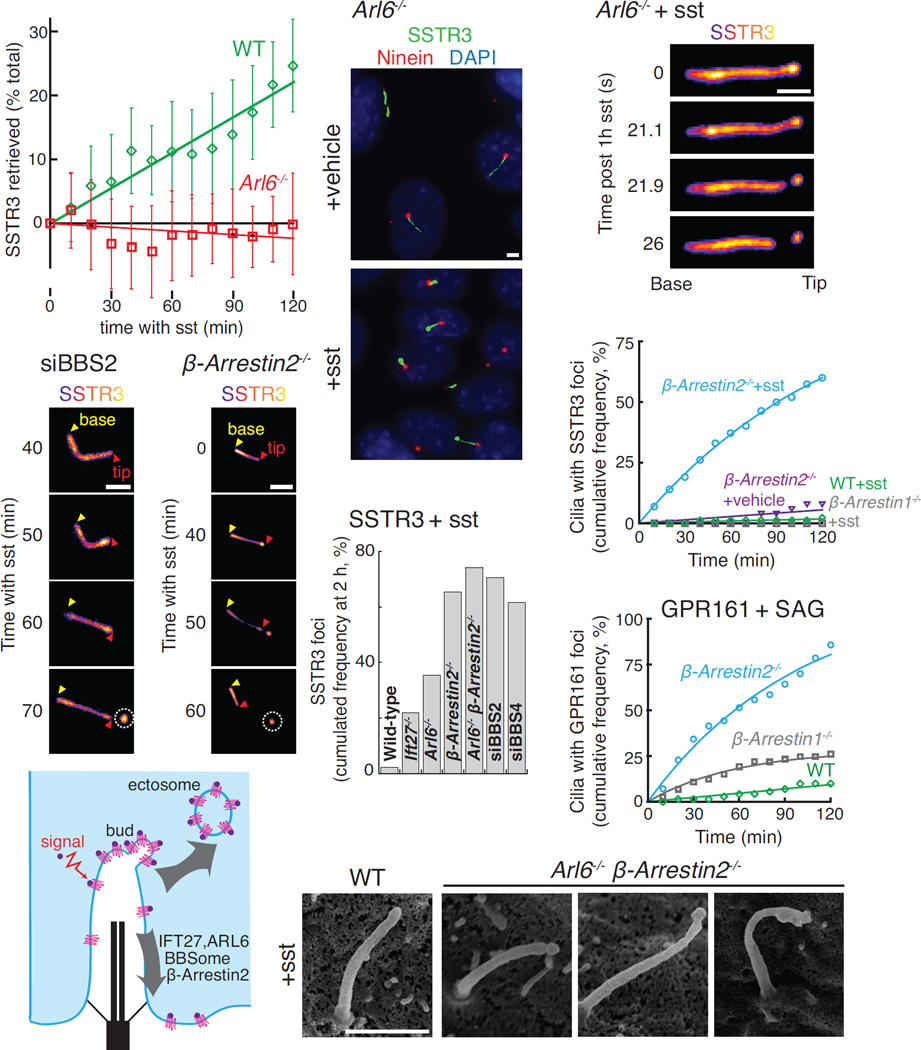
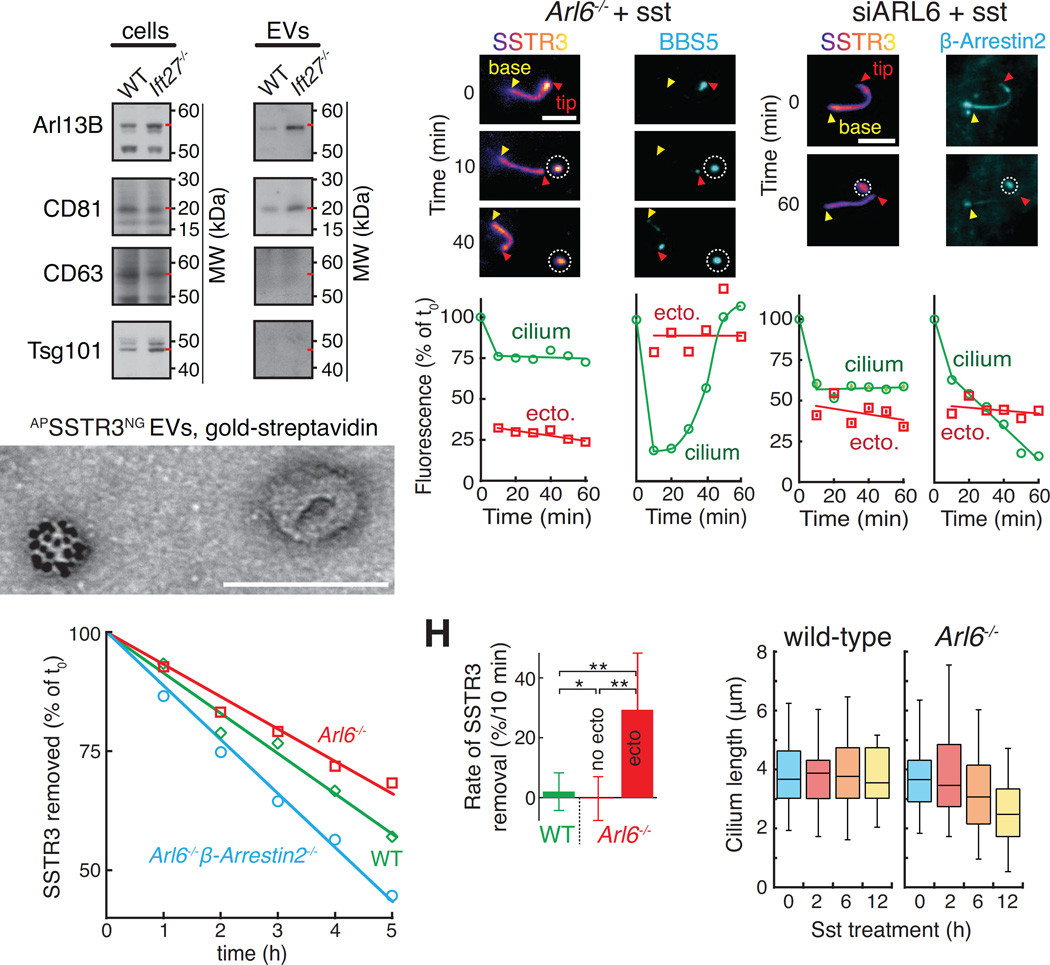
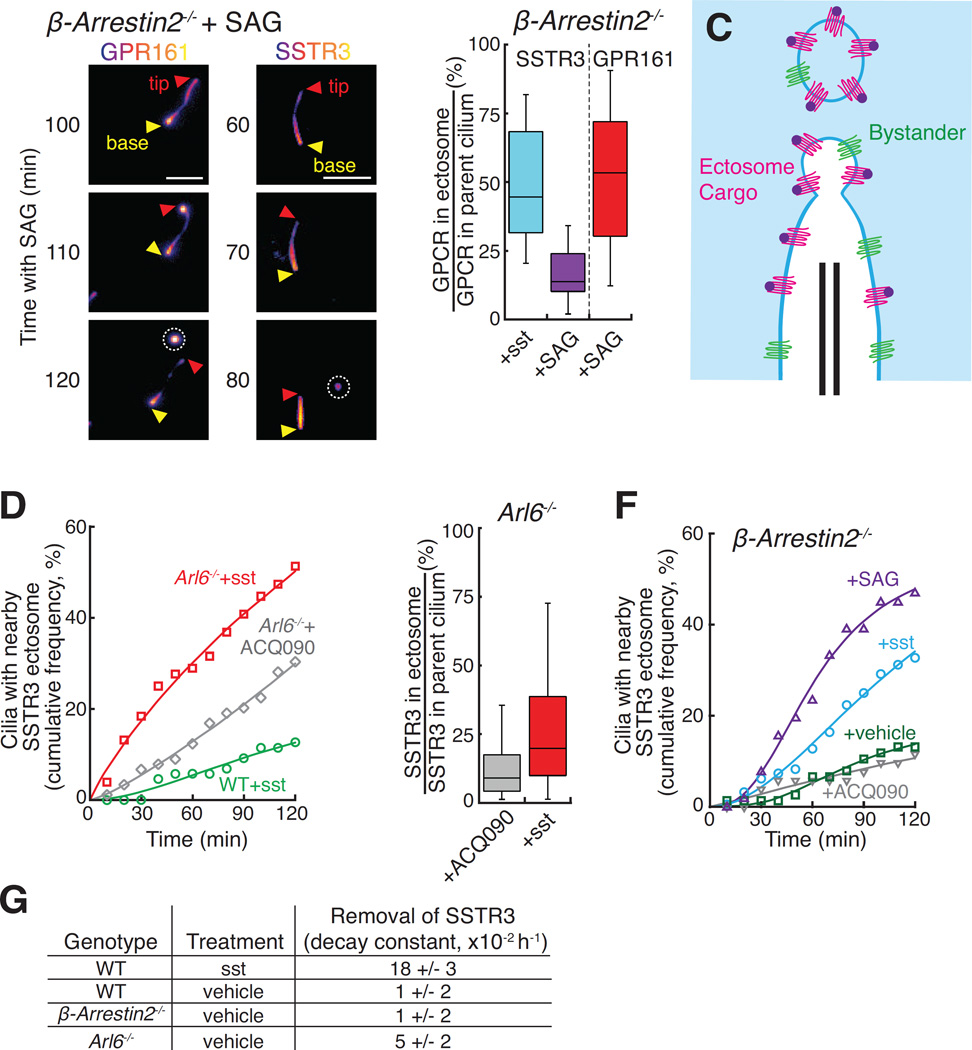
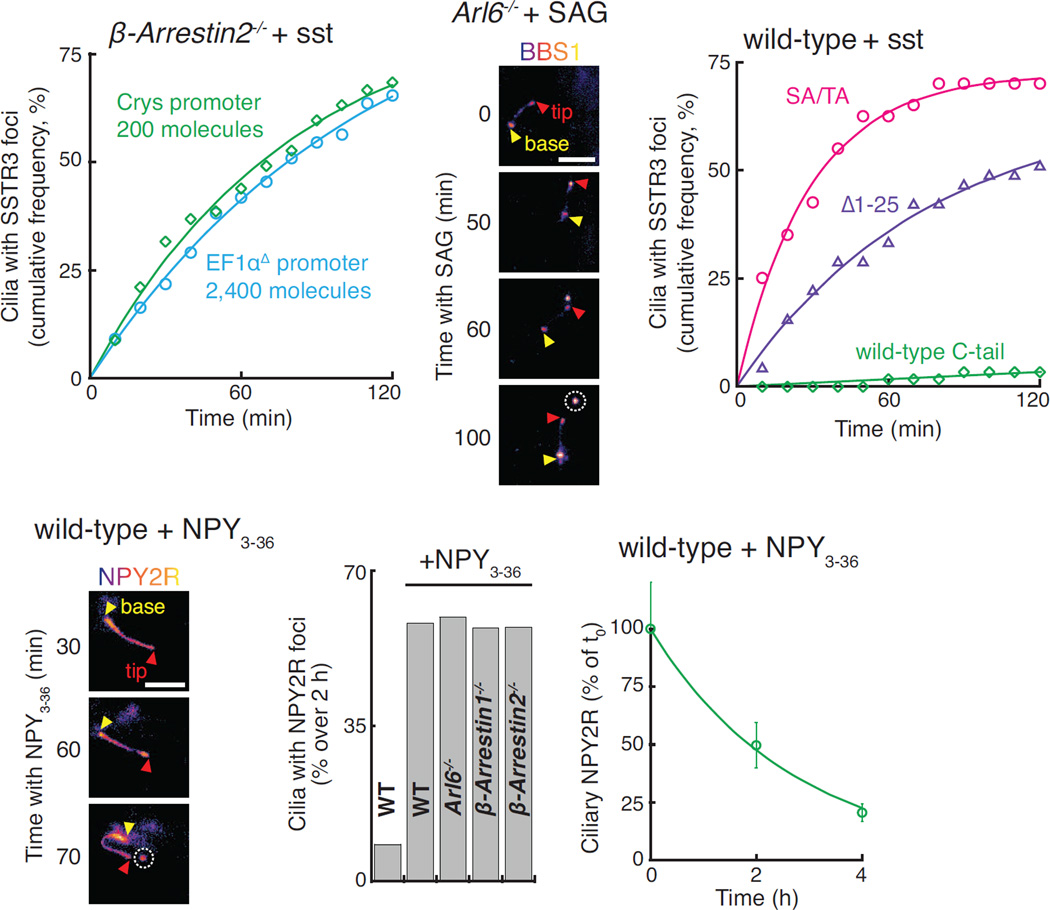
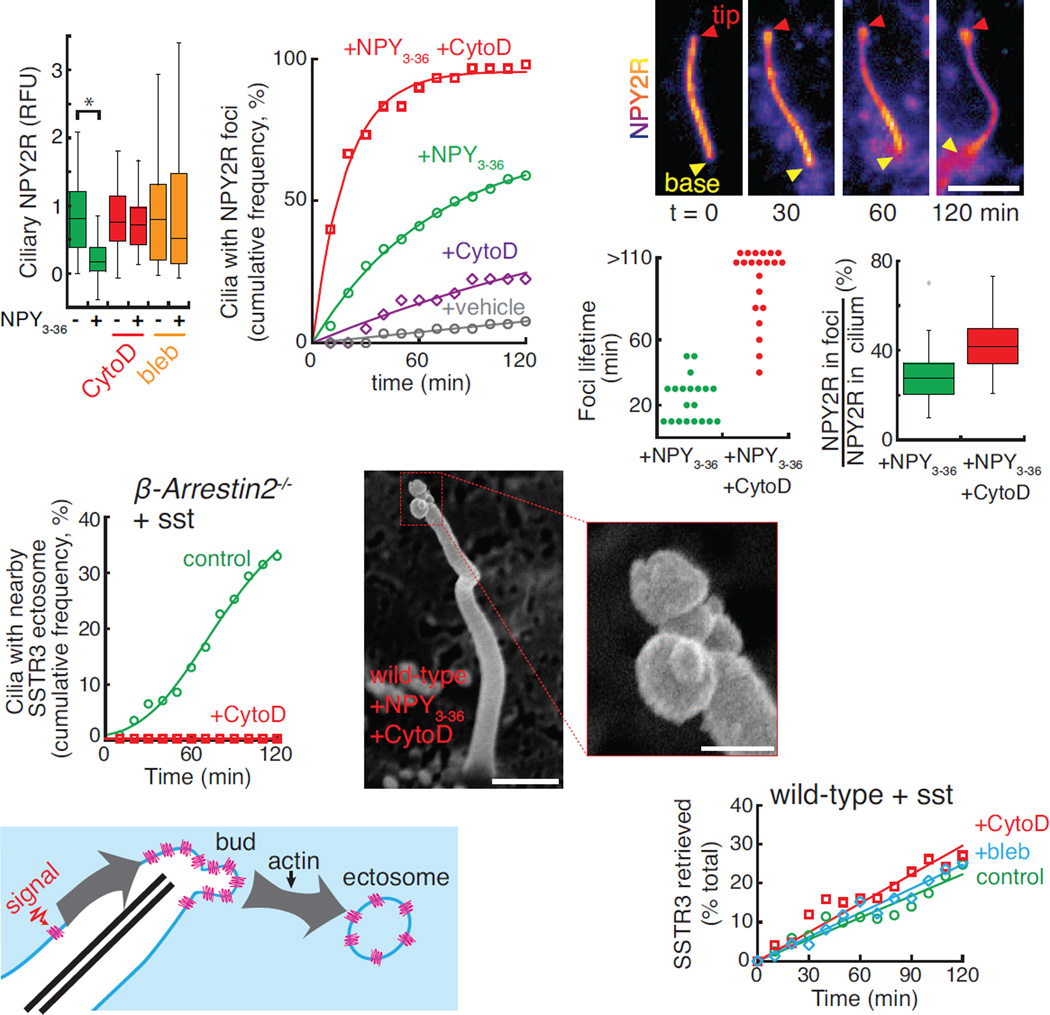
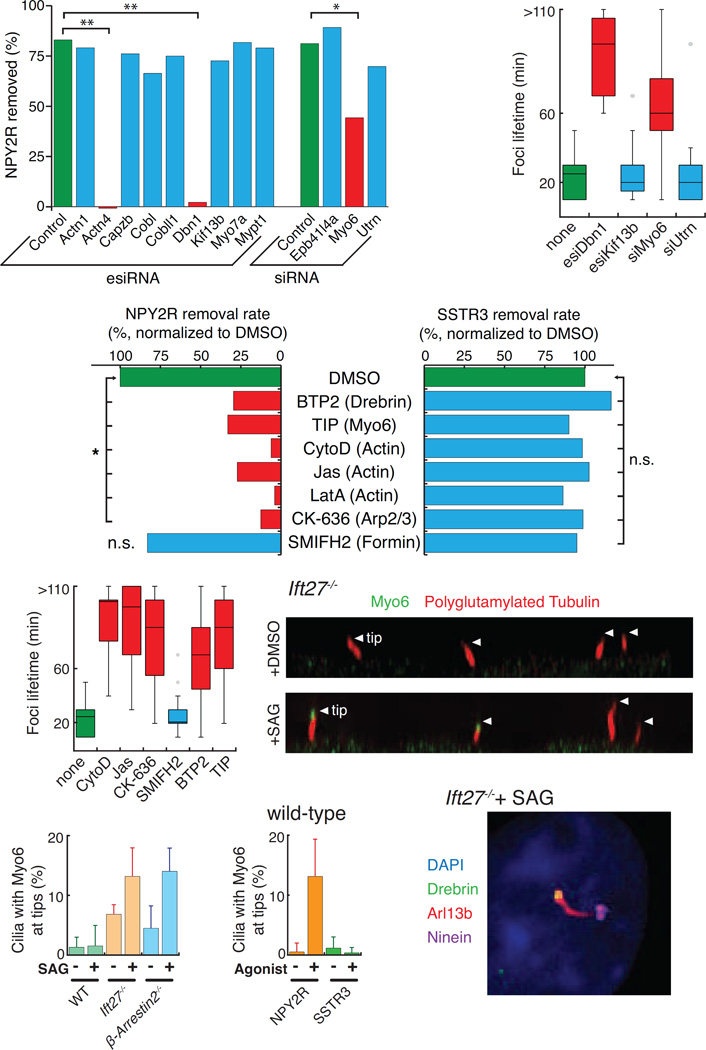
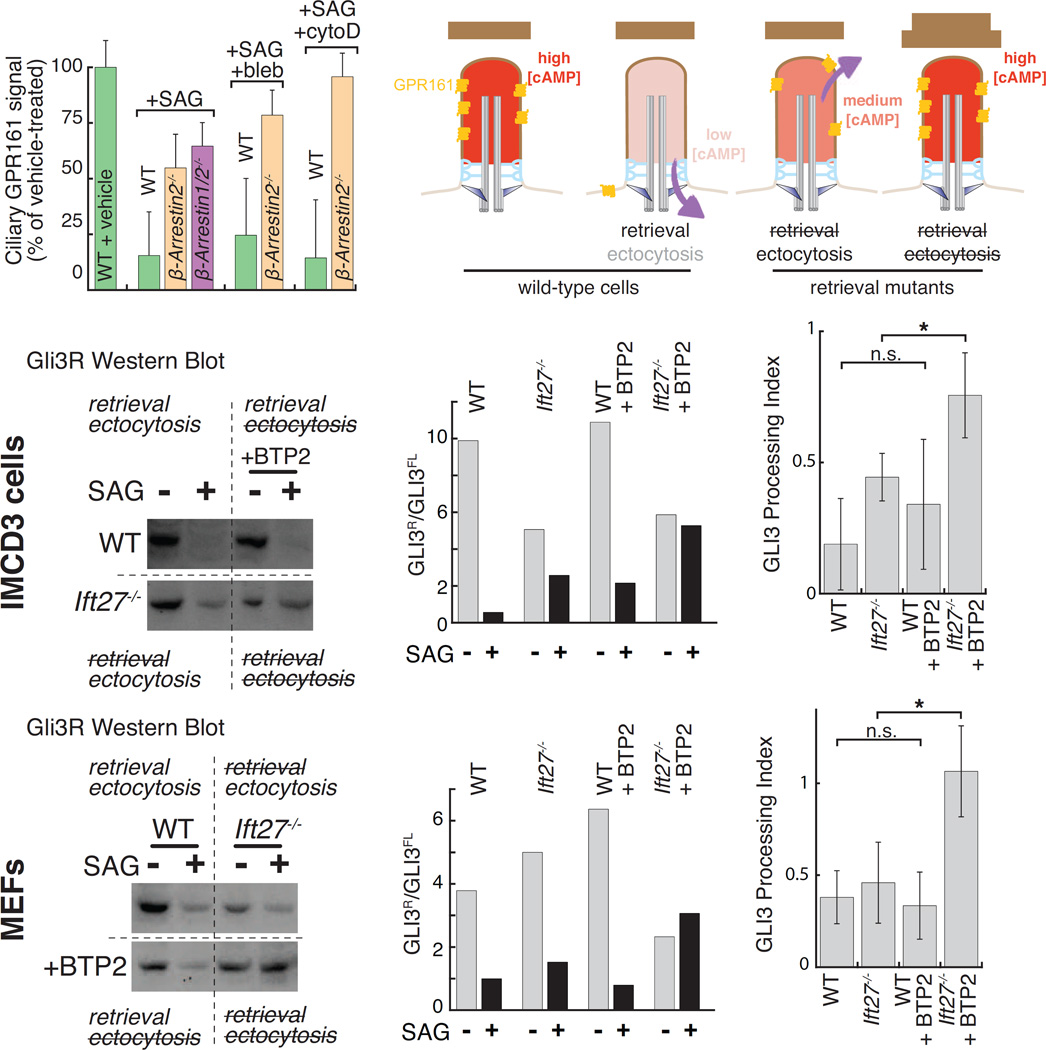
Signaling receptors dynamically exit cilia upon activation of signaling pathways such as Hedgehog. Here, we find that when activated G protein-coupled receptors (GPCRs) fail to undergo BBSome-mediated retrieval from cilia back into the cell, these GPCRs concentrate into membranous buds at the tips of cilia before release into extracellular vesicles named ectosomes. Unexpectedly, actin and the actin regulators drebrin and myosin 6 mediate ectosome release from the tip of cilia. Mirroring signal-dependent retrieval, signal-dependent ectocytosis is a selective and effective process that removes activated signaling molecules from cilia. Congruently, ectocytosis compensates for BBSome defects as ectocytic removal of GPR161, a negative regulator of Hedgehog signaling, permits the appropriate transduction of Hedgehog signals in Bbs mutants. Finally, ciliary receptors that lack retrieval determinants such as the anorexigenic GPCR NPY2R undergo signal-dependent ectocytosis in wild-type cells. Our data show that signal-dependent ectocytosis regulates ciliary signaling in physiological and pathological contexts.
Keywords: BBSome; GPCR; Hedgehog; actin; cilia; drebrin; exosomes; extracellular vesicles; myosin 6.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Barlowe C, Helenius A. Cargo Capture and Bulk Flow in the Early Secretory Pathway. Annu Rev Cell Dev Biol. 2016;32:197–222. - PubMed
-
- Buss F, Kendrick-Jones J, Lionne C, Knight AE, Côté GP, Paul Luzio J. The localization of myosin VI at the golgi complex and leading edge of fibroblasts and its phosphorylation and recruitment into membrane ruffles of A431 cells after growth factor stimulation. J Cell Biol. 1998;143:1535–1545. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
