

De Novo Mutations in EBF3 Cause a Neurodevelopmental Syndrome
- PMID: 28017370
- PMCID: PMC5223060
- DOI: 10.1016/j.ajhg.2016.11.020
De Novo Mutations in EBF3 Cause a Neurodevelopmental Syndrome
Abstract
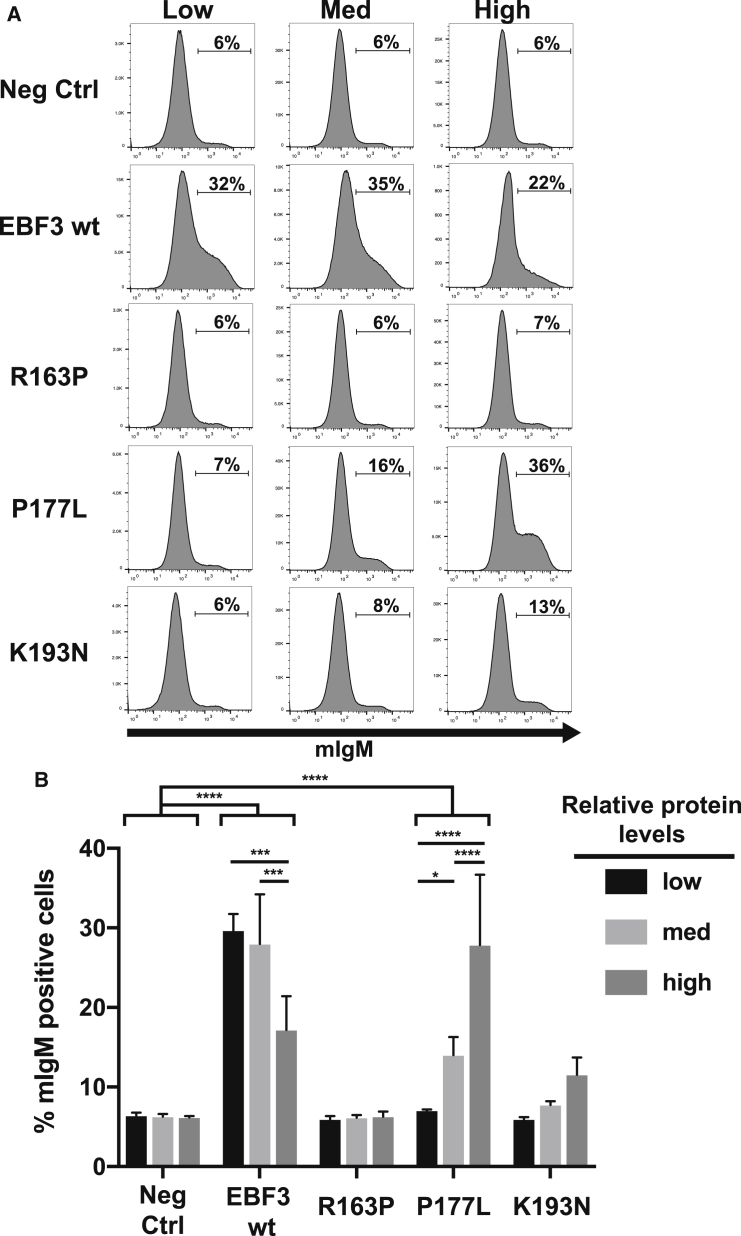
Early B cell factor 3 (EBF3) is an atypical transcription factor that is thought to influence the laminar formation of the cerebral cortex. Here, we report that de novo mutations in EBF3 cause a complex neurodevelopmental syndrome. The mutations were identified in two large-scale sequencing projects: the UK Deciphering Developmental Disorders (DDD) study and the Canadian Clinical Assessment of the Utility of Sequencing and Evaluation as a Service (CAUSES) study. The core phenotype includes moderate to severe intellectual disability, and many individuals exhibit cerebellar ataxia, subtle facial dysmorphism, strabismus, and vesicoureteric reflux, suggesting that EBF3 has a widespread developmental role. Pathogenic de novo variants identified in EBF3 include multiple loss-of-function and missense mutations. Structural modeling suggested that the missense mutations affect DNA binding. Functional analysis of mutant proteins with missense substitutions revealed reduced transcriptional activities and abilities to form heterodimers with wild-type EBF3. We conclude that EBF3, a transcription factor previously unknown to be associated with human disease, is important for brain and other organ development and warrants further investigation.
Copyright © 2017 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Mutations in EBF3 Disturb Transcriptional Profiles and Cause Intellectual Disability, Ataxia, and Facial Dysmorphism.Am J Hum Genet. 2017 Jan 5;100(1):117-127. doi: 10.1016/j.ajhg.2016.11.012. Epub 2016 Dec 22. Am J Hum Genet. 2017. PMID: 28017373 Free PMC article.
-
A Syndromic Neurodevelopmental Disorder Caused by De Novo Variants in EBF3.Am J Hum Genet. 2017 Jan 5;100(1):128-137. doi: 10.1016/j.ajhg.2016.11.018. Epub 2016 Dec 22. Am J Hum Genet. 2017. PMID: 28017372 Free PMC article.
-
Novel de novo variant in EBF3 is likely to impact DNA binding in a patient with a neurodevelopmental disorder and expanded phenotypes: patient report, in silico functional assessment, and review of published cases.Cold Spring Harb Mol Case Stud. 2017 May;3(3):a001743. doi: 10.1101/mcs.a001743. Cold Spring Harb Mol Case Stud. 2017. PMID: 28487885 Free PMC article.
-
A nonsense variant in the C-terminal transactivation domain of the EBF3 gene in an individual with intellectual disability and behavioural disorder: case report and literature review.Psychiatr Genet. 2025 Jun 1;35(3):75-78. doi: 10.1097/YPG.0000000000000386. Epub 2025 Feb 28. Psychiatr Genet. 2025. PMID: 40073166 Review.
-
PURA syndrome: clinical delineation and genotype-phenotype study in 32 individuals with review of published literature.J Med Genet. 2018 Feb;55(2):104-113. doi: 10.1136/jmedgenet-2017-104946. Epub 2017 Nov 2. J Med Genet. 2018. PMID: 29097605 Free PMC article. Review.
Cited by
-
Generation and Characterization of a Knockout Mouse of an Enhancer of EBF3.bioRxiv [Preprint]. 2025 Jan 10:2025.01.09.631762. doi: 10.1101/2025.01.09.631762. bioRxiv. 2025. PMID: 39829799 Free PMC article. Preprint.
-
Open chromatin dynamics in prosensory cells of the embryonic mouse cochlea.Sci Rep. 2019 Jun 21;9(1):9060. doi: 10.1038/s41598-019-45515-2. Sci Rep. 2019. PMID: 31227770 Free PMC article.
-
Coding and noncoding variants in EBF3 are involved in HADDS and simplex autism.Hum Genomics. 2021 Jul 13;15(1):44. doi: 10.1186/s40246-021-00342-3. Hum Genomics. 2021. PMID: 34256850 Free PMC article.
-
Disrupted development of sensory systems and the cerebellum in a zebrafish ebf3a mutant.G3 (Bethesda). 2025 Jul 9;15(7):jkaf115. doi: 10.1093/g3journal/jkaf115. G3 (Bethesda). 2025. PMID: 40406967 Free PMC article.
-
Neuroimaging Findings in Patients with EBF3 Mutations: Report of Two Cases.Mol Syndromol. 2021 Jun;12(3):186-193. doi: 10.1159/000513583. Epub 2021 Apr 9. Mol Syndromol. 2021. PMID: 34177436 Free PMC article.
References
-
- Firth H.V., Wright C.F., DDD Study The Deciphering Developmental Disorders (DDD) study. Dev. Med. Child Neurol. 2011;53:702–703. - PubMed
-
- Wright C.F., Fitzgerald T.W., Jones W.D., Clayton S., McRae J.F., van Kogelenberg M., King D.A., Ambridge K., Barrett D.M., Bayzetinova T., DDD study Genetic diagnosis of developmental disorders in the DDD study: a scalable analysis of genome-wide research data. Lancet. 2015;385:1305–1314. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
