

Autosomal Recessive Keratoderma-Ichthyosis-Deafness (ARKID) Syndrome Is Caused by VPS33B Mutations Affecting Rab Protein Interaction and Collagen Modification
- PMID: 28017832
- PMCID: PMC5358661
- DOI: 10.1016/j.jid.2016.12.010
Autosomal Recessive Keratoderma-Ichthyosis-Deafness (ARKID) Syndrome Is Caused by VPS33B Mutations Affecting Rab Protein Interaction and Collagen Modification
Abstract
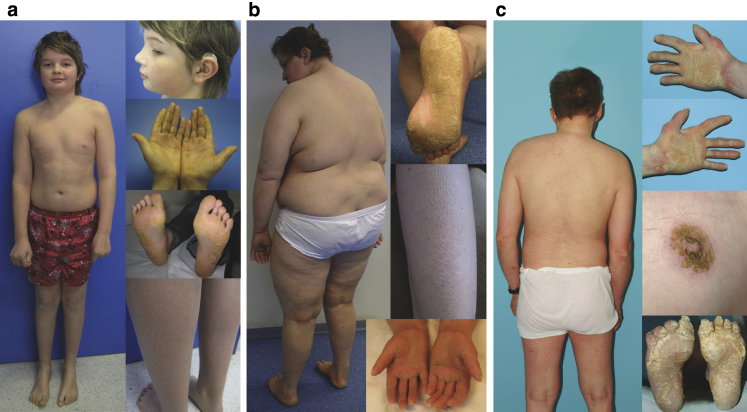
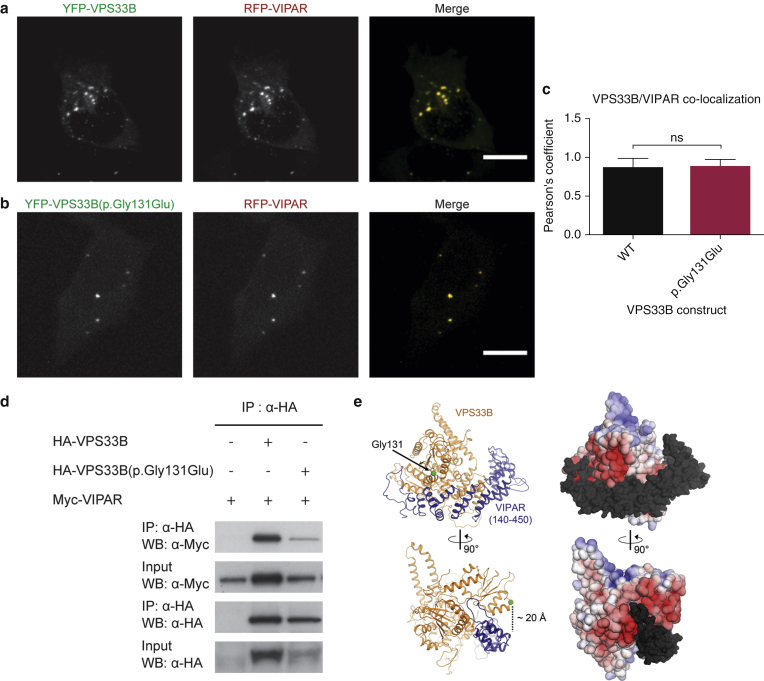
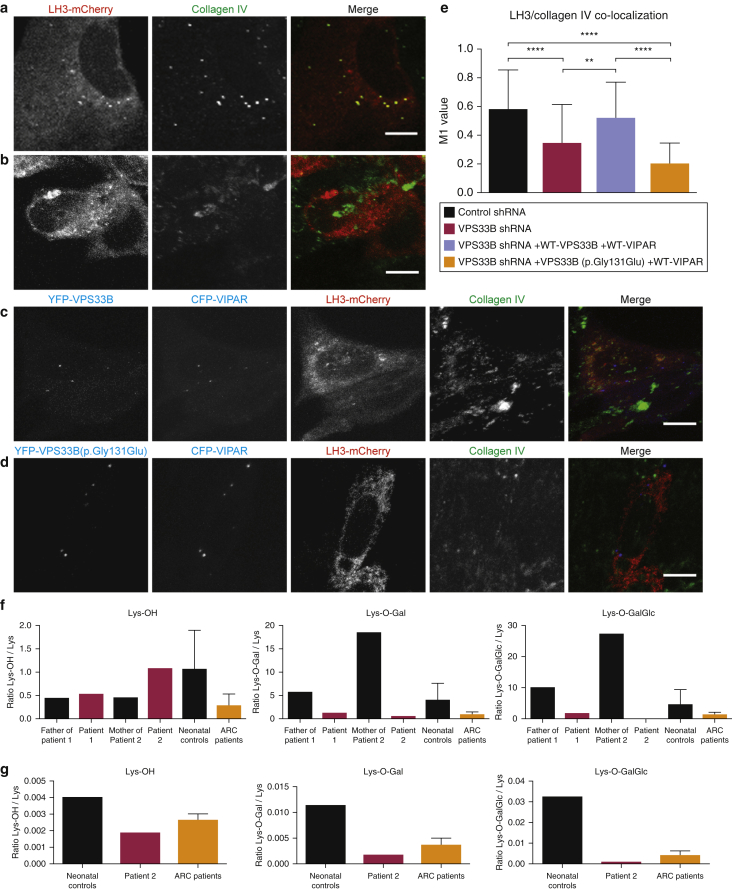
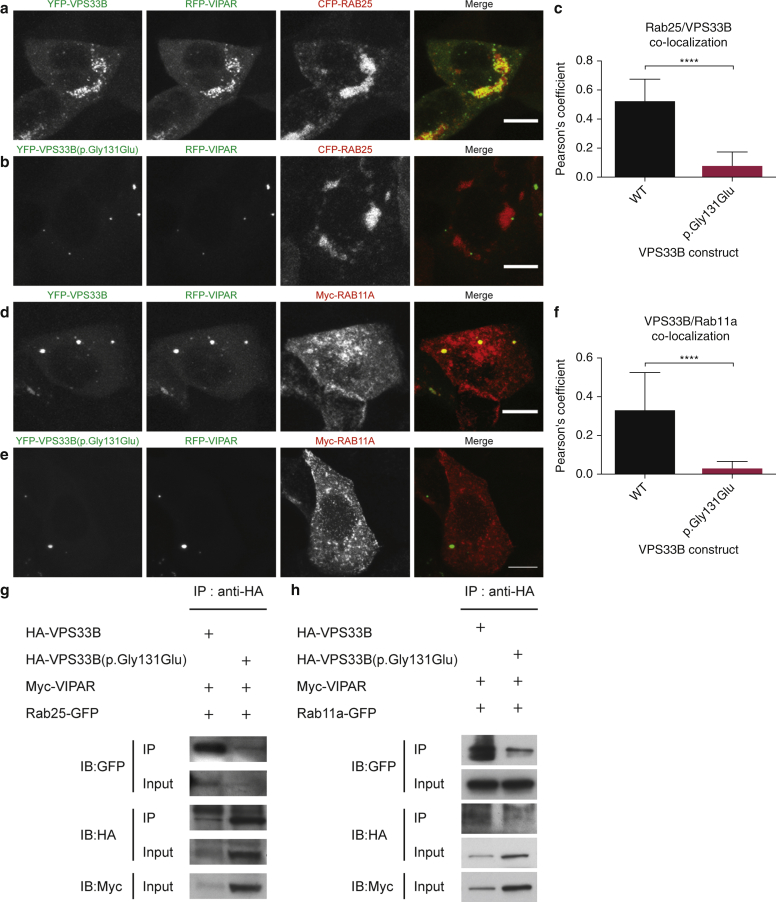
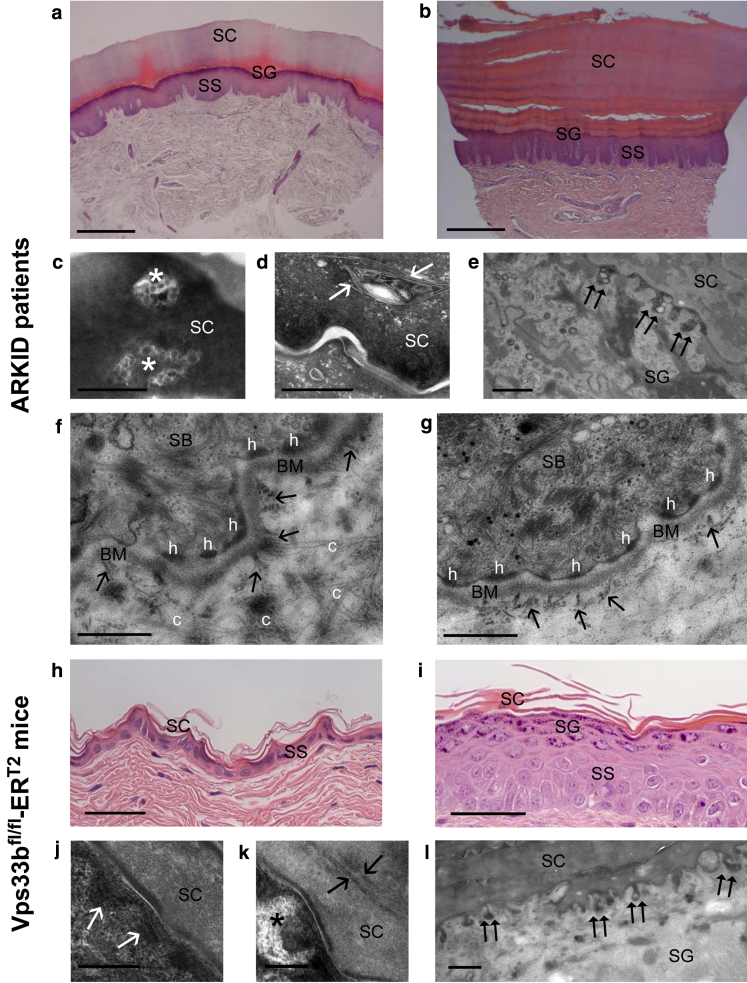
In this paper, we report three patients with severe palmoplantar keratoderma associated with ichthyosis and sensorineural deafness. Biallelic mutations were found in VPS33B, encoding VPS33B, a Sec1/Munc18 family protein that interacts with Rab11a and Rab25 proteins and is involved in trafficking of the collagen-modifying enzyme LH3. Two patients were homozygous for the missense variant p.Gly131Glu, whereas one patient was compound heterozygous for p.Gly131Glu and the splice site mutation c.240-1G>C, previously reported in patients with arthrogryposis renal dysfunction and cholestasis syndrome. We demonstrated the pathogenicity of variant p.Gly131Glu by assessing the interactions of the mutant VPS33B construct and its ability to traffic LH3. Compared with wild-type VPS33B, the p.Gly131Glu mutant VPS33B had reduced coimmunoprecipitation and colocalization with Rab11a and Rab25 and did not rescue LH3 trafficking. Confirming the cell-based experiments, we found deficient LH3-specific collagen lysine modifications in patients' urine and skin fibroblasts. Additionally, the epidermal ultrastructure of the p.Gly131Glu patients mirrored defects in tamoxifen-inducible VPS33B-deficient Vps33bfl/fl-ERT2 mice. Both patients and murine models revealed an impaired epidermal structure, ascribed to aberrant secretion of lamellar bodies, which are essential for epidermal barrier formation. Our results demonstrate that p.Gly131Glu mutant VPS33B causes an autosomal recessive keratoderma-ichthyosis-deafness syndrome.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Abu-Sa’da O., Barbar M., Al-Harbi N., Taha D. Arthrogryposis, renal tubular acidosis and cholestasis (ARC) syndrome: two new cases and review. Clin Dysmorphol. 2005;14:191–196. - PubMed
-
- Balderhaar H.J., Ungermann C. CORVET and HOPS tethering complexes—coordinators of endosome and lysosome fusion. J Cell Sci. 2013;126:1307–1316. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
