

The Calcium-Sensing Receptor and the Parathyroid: Past, Present, Future
- PMID: 28018229
- PMCID: PMC5156698
- DOI: 10.3389/fphys.2016.00563
The Calcium-Sensing Receptor and the Parathyroid: Past, Present, Future
Abstract
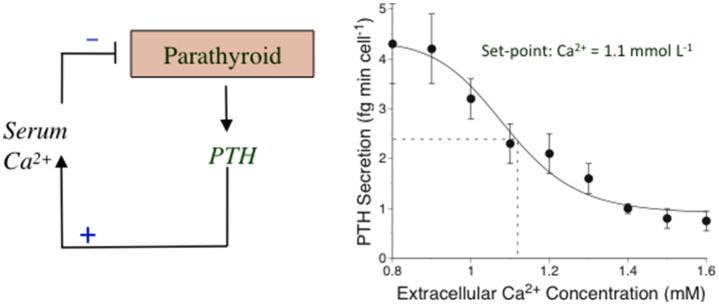
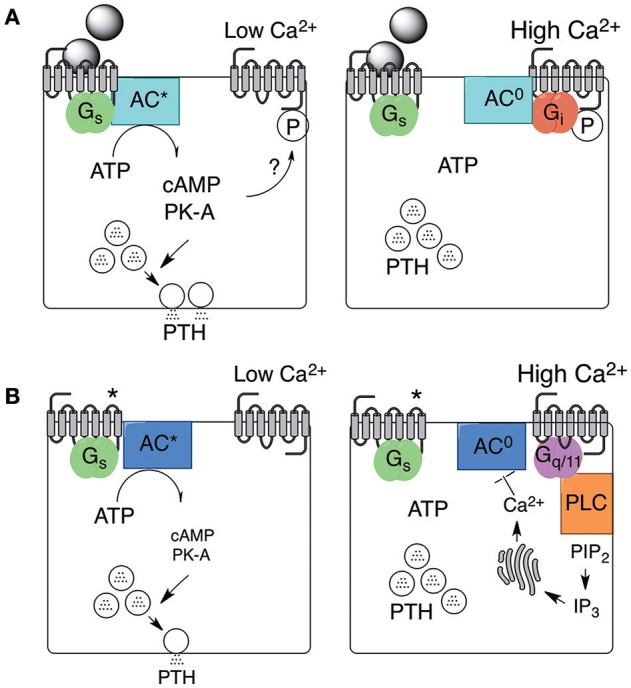
Parathyroid hormone (PTH) defends the extracellular fluid from hypocalcemia and has powerful and well-documented actions on the skeleton and renal tubular system. To achieve a satisfactory stable plasma calcium level, the secretion of PTH, and the resulting serum PTH level, is titrated carefully to the prevailing plasma ionized Ca2+ concentration via a Ca2+ sensing mechanism that mediates feedback inhibition of PTH secretion. Herein, I consider the properties of the parathyroid Ca2+ sensing mechanism, the identity of the Ca2+ sensor, the intracellular biochemical mechanisms that it controls, the manner of its integration with other components of the PTH secretion control mechanism, and its modulation by other nutrients. Together the well-established, recently elucidated, and yet-to-be discovered elements of the story constitute the past, present, and future of the parathyroid and its calcium-sensing receptor (CaSR).
Keywords: Calcimimetics; adenylate cyclase; calcilytics; calcium-sensing receptor; heterotrimeric G proteins; mineral metabolism; parathyroid; phospholipase C.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous