

Analysis of the Interaction between Globular Head Modules of Human C1q and Its Candidate Receptor gC1qR
- PMID: 28018340
- PMCID: PMC5153404
- DOI: 10.3389/fimmu.2016.00567
Analysis of the Interaction between Globular Head Modules of Human C1q and Its Candidate Receptor gC1qR
Abstract
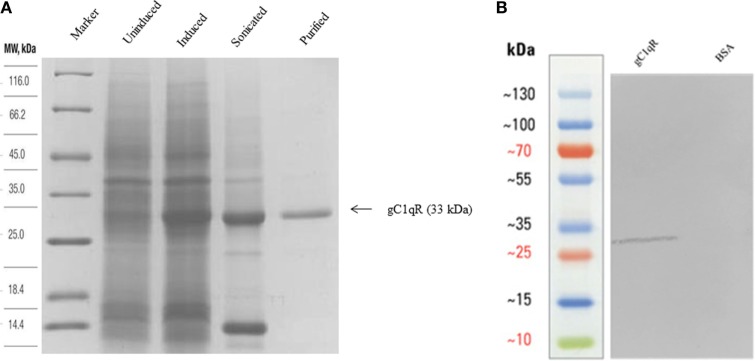
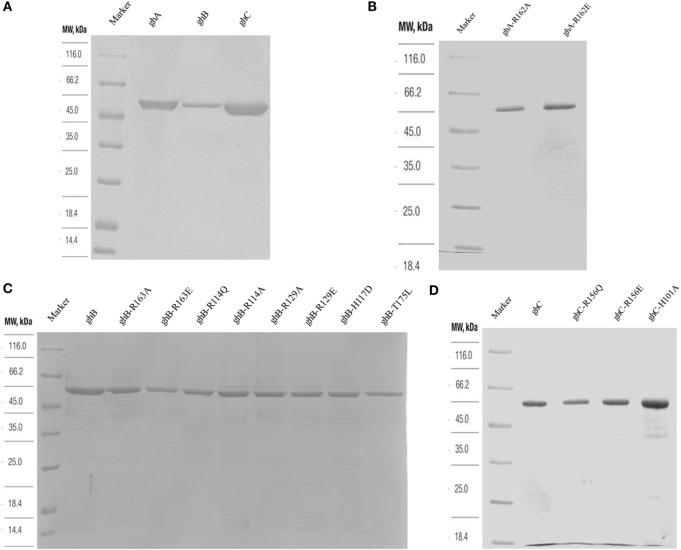
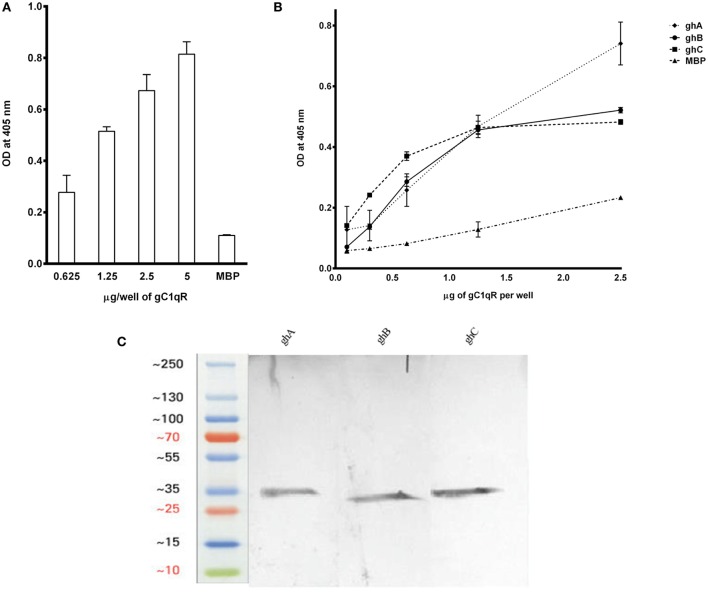
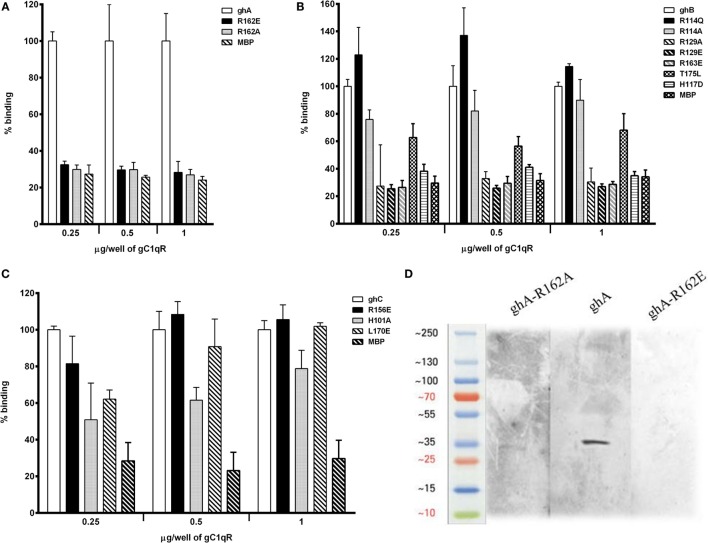
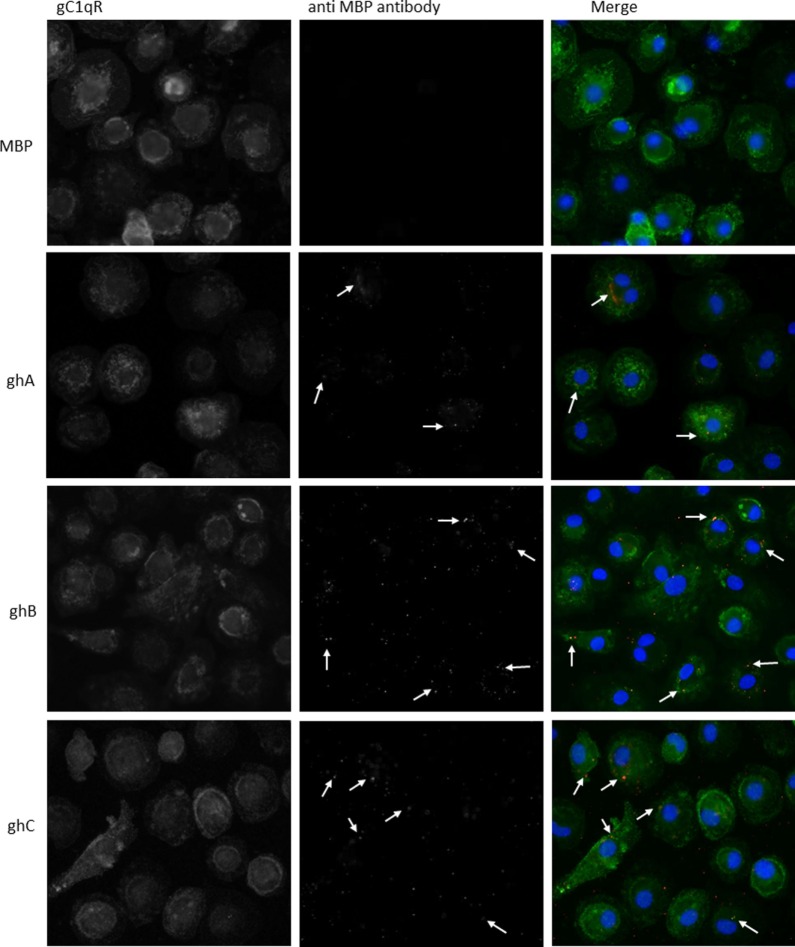
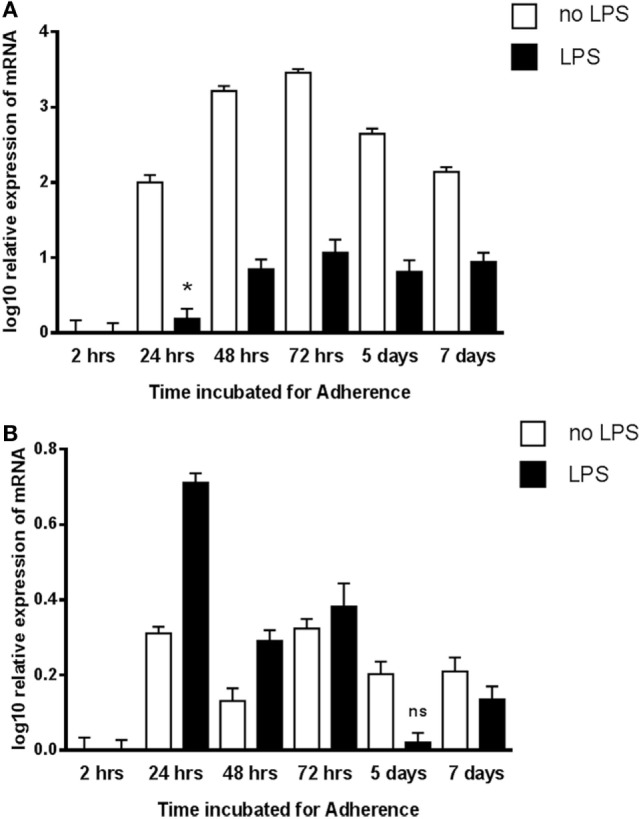
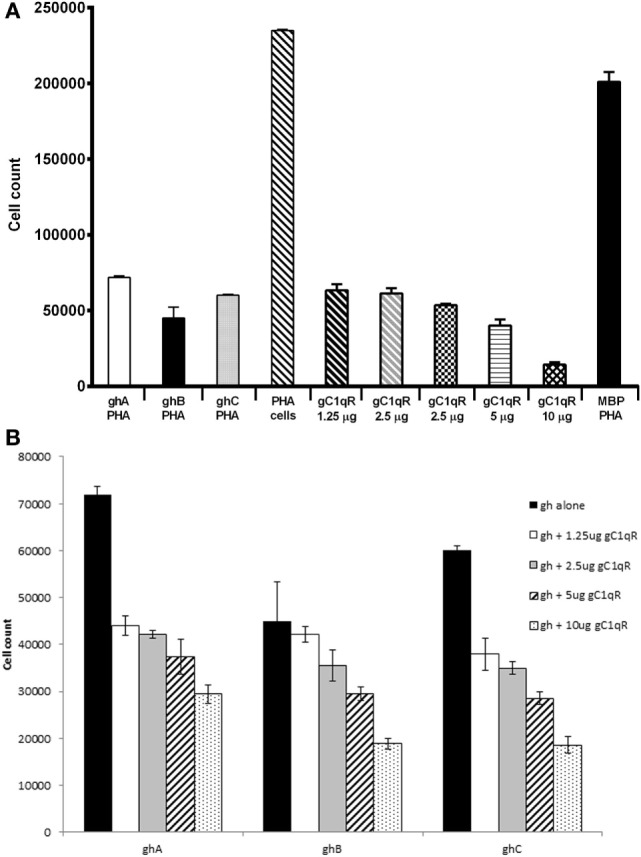
The heterotrimeric globular head (gC1q) domain of human C1q is made up of the C-terminal ends of the three individual chains, ghA, ghB, and ghC. A candidate receptor for the gC1q domain is a multi-functional pattern recognition protein, gC1qR. Since understanding of gC1qR and gC1q interaction could provide an insight into the pleiotropic functions of gC1qR, this study was undertaken to identify the gC1qR-binding site on the gC1q domain, using the recombinant ghA, ghB, and ghC modules and their substitution mutants. Our results show that ghA, ghB, and ghC modules can interact with gC1qR independently, thus reinforcing the notion of modularity within the gC1q domain of human C1q. Mutational analysis revealed that while Arg162 in the ghA module is central to interaction between gC1qR and C1q, a single amino acid substitution (arginine to glutamate) in residue 114 of the ghB module resulted in enhanced binding. Expression of gC1qR and C1q in adherent monocytes with or without pro-inflammatory stimuli was also analyzed by qPCR; it showed an autocrine/paracrine basis of C1q and gC1qR interaction. Microscopic studies revealed that C1q and gC1qR are colocalized on PBMCs. Cell proliferation assays indicated that ghA, ghB, and ghC modules were able to attenuate phytohemagglutinin-stimulated proliferation of PBMCs. Addition of gC1qR had an additive effect on the anti-proliferative effect of globular head modules. In summary, our results identify residues involved in C1q-gC1qR interaction and explain, to a certain level, their involvement on the immune cell surface, which is relevant for C1q-induced functions including inflammation, infection, and immunity.
Keywords: C1q; cell proliferation; gC1qR; globular head; protein–protein interaction.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases