

Roles of Aquaporins in Setaria viridis Stem Development and Sugar Storage
- PMID: 28018372
- PMCID: PMC5147461
- DOI: 10.3389/fpls.2016.01815
Roles of Aquaporins in Setaria viridis Stem Development and Sugar Storage
Abstract
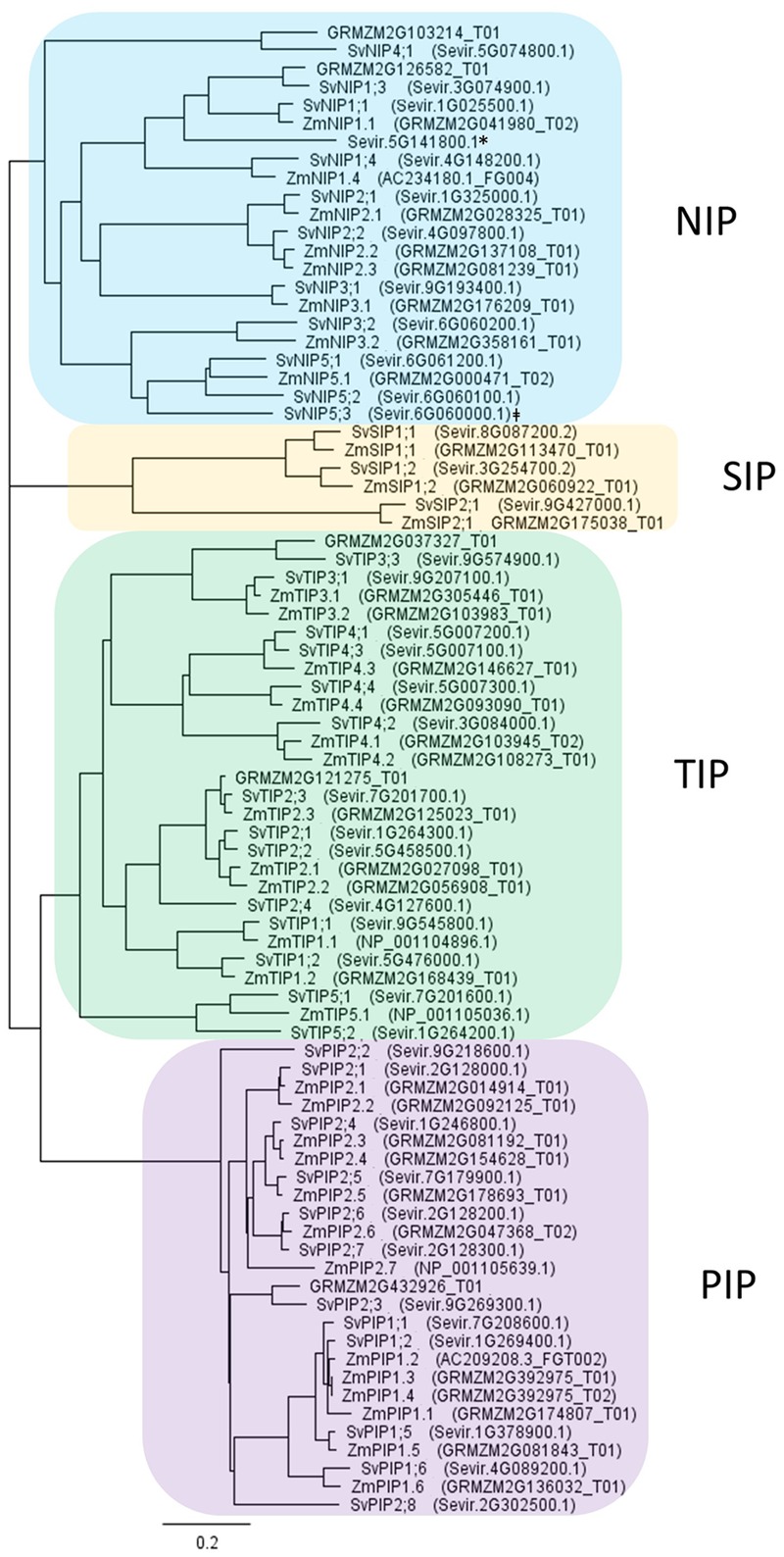
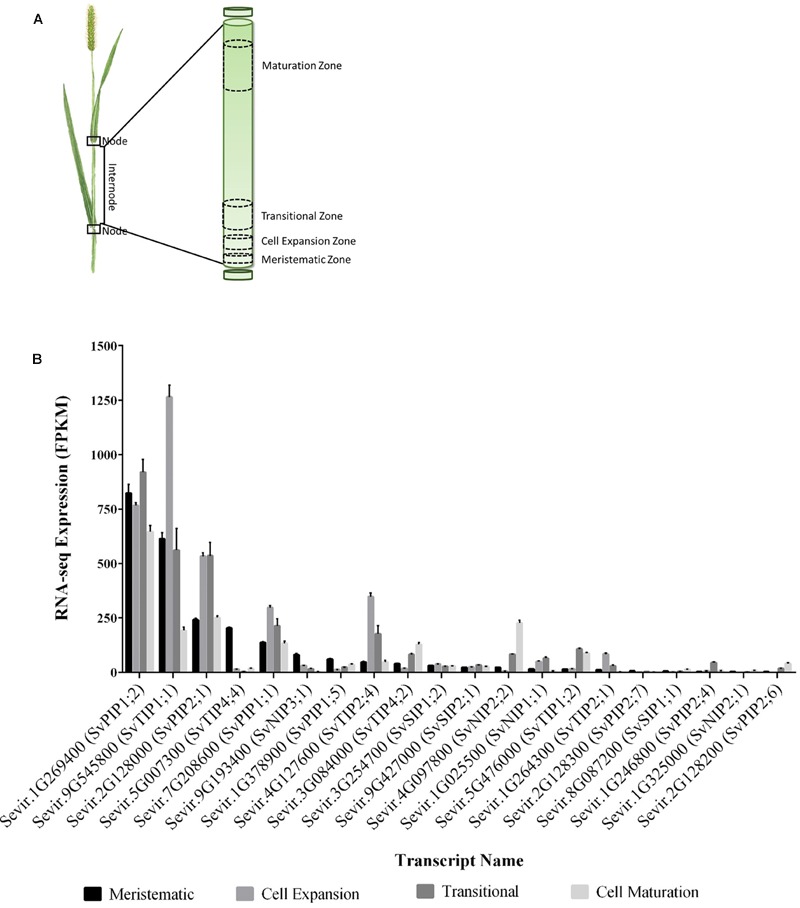
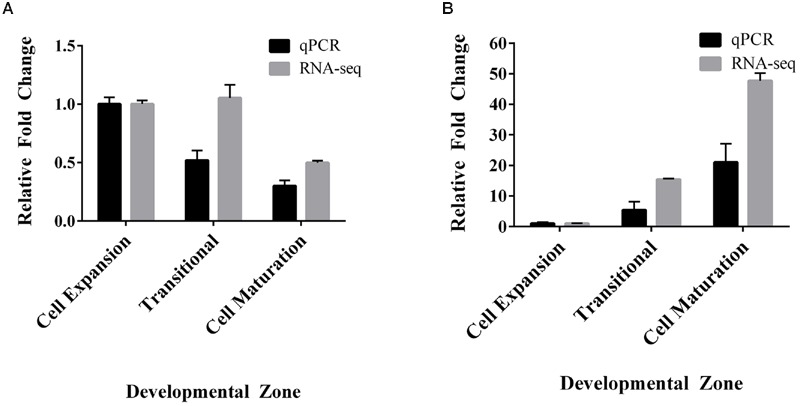
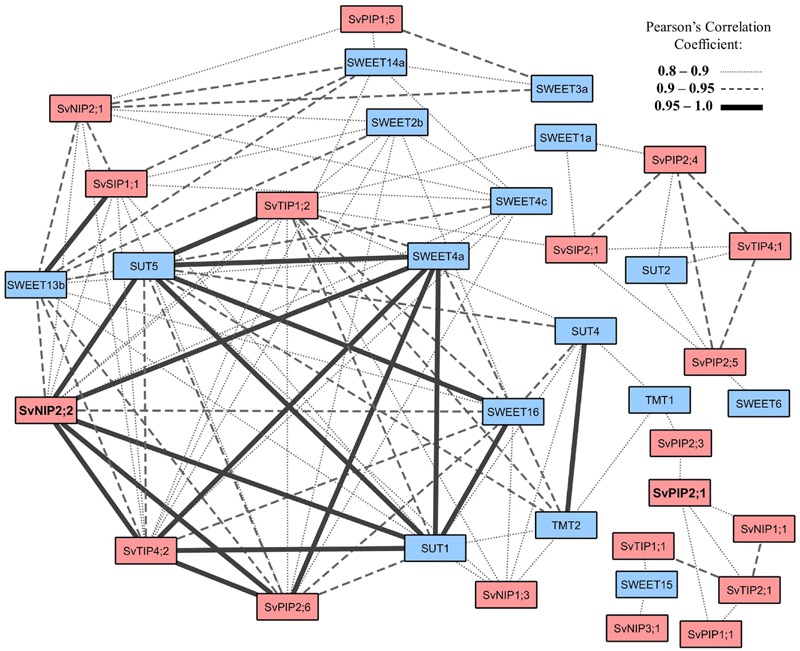
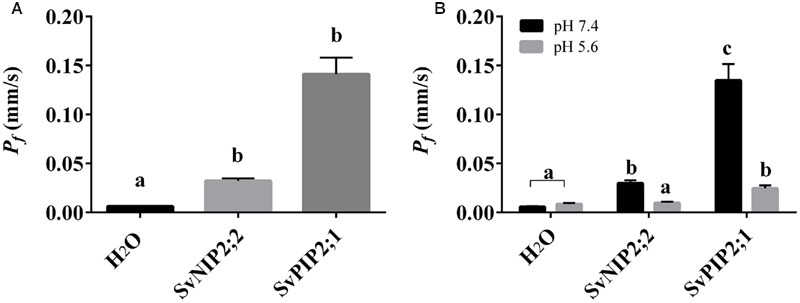
Setaria viridis is a C4 grass used as a model for bioenergy feedstocks. The elongating internodes in developing S. viridis stems grow from an intercalary meristem at the base, and progress acropetally toward fully expanded cells that store sugar. During stem development and maturation, water flow is a driver of cell expansion and sugar delivery. As aquaporin proteins are implicated in regulating water flow, we analyzed elongating and mature internode transcriptomes to identify putative aquaporin encoding genes that had particularly high transcript levels during the distinct stages of internode cell expansion and maturation. We observed that SvPIP2;1 was highly expressed in internode regions undergoing cell expansion, and SvNIP2;2 was highly expressed in mature sugar accumulating regions. Gene co-expression analysis revealed SvNIP2;2 expression was highly correlated with the expression of five putative sugar transporters expressed in the S. viridis internode. To explore the function of the proteins encoded by SvPIP2;1 and SvNIP2;2, we expressed them in Xenopus laevis oocytes and tested their permeability to water. SvPIP2;1 and SvNIP2;2 functioned as water channels in X. laevis oocytes and their permeability was gated by pH. Our results indicate that SvPIP2;1 may function as a water channel in developing stems undergoing cell expansion and SvNIP2;2 is a candidate for retrieving water and possibly a yet to be determined solute from mature internodes. Future research will investigate whether changing the function of these proteins influences stem growth and sugar yield in S. viridis.
Keywords: aquaporin; grasses; stem; sugar accumulation; water transport.
Figures
Similar articles
-
A developing Setaria viridis internode: an experimental system for the study of biomass generation in a C4 model species.Biotechnol Biofuels. 2016 Feb 24;9:45. doi: 10.1186/s13068-016-0457-6. eCollection 2016. Biotechnol Biofuels. 2016. PMID: 26918029 Free PMC article.
-
Comprehensive expression analyses of the ABCG subfamily reveal SvABCG17 as a potential transporter of lignin monomers in the model C4 grass Setaria viridis.J Plant Physiol. 2023 Jan;280:153900. doi: 10.1016/j.jplph.2022.153900. Epub 2022 Dec 13. J Plant Physiol. 2023. PMID: 36525838
-
Elucidating the role of SWEET13 in phloem loading of the C4 grass Setaria viridis.Plant J. 2022 Feb;109(3):615-632. doi: 10.1111/tpj.15581. Epub 2021 Dec 12. Plant J. 2022. PMID: 34780111
-
Discovery of aquaporins: a breakthrough in research on renal water transport.Pediatr Nephrol. 1995 Apr;9(2):228-34. doi: 10.1007/BF00860757. Pediatr Nephrol. 1995. PMID: 7540850 Review.
-
Setaria viridis and Setaria italica, model genetic systems for the Panicoid grasses.J Exp Bot. 2011 May;62(9):3031-7. doi: 10.1093/jxb/err096. Epub 2011 Mar 31. J Exp Bot. 2011. PMID: 21459768 Review.
Cited by
-
Multi-omics intervention in Setaria to dissect climate-resilient traits: Progress and prospects.Front Plant Sci. 2022 Aug 31;13:892736. doi: 10.3389/fpls.2022.892736. eCollection 2022. Front Plant Sci. 2022. PMID: 36119586 Free PMC article. Review.
-
Study on aquaporins of Setaria italica suggests the involvement of SiPIP3;1 and SiSIP1;1 in abiotic stress response.Funct Integr Genomics. 2019 Jul;19(4):587-596. doi: 10.1007/s10142-018-00653-0. Epub 2019 Feb 13. Funct Integr Genomics. 2019. PMID: 30759293
-
Genome-Wide Identification and Functional Characterization of SKP1-like Gene Family Reveal Its Involvement in Response to Stress in Cotton.Int J Mol Sci. 2025 Jan 6;26(1):418. doi: 10.3390/ijms26010418. Int J Mol Sci. 2025. PMID: 39796275 Free PMC article.
-
Structure, evolution and diverse physiological roles of SWEET sugar transporters in plants.Plant Mol Biol. 2019 Jul;100(4-5):351-365. doi: 10.1007/s11103-019-00872-4. Epub 2019 Apr 27. Plant Mol Biol. 2019. PMID: 31030374 Review.
-
Whole-Genome and Expression Analyses of Bamboo Aquaporin Genes Reveal Their Functions Involved in Maintaining Diurnal Water Balance in Bamboo Shoots.Cells. 2018 Nov 2;7(11):195. doi: 10.3390/cells7110195. Cells. 2018. PMID: 30400256 Free PMC article.
References
-
- Azad A. K., Ahmed J., Alum A., Hasan M., Ishikawa T., Sawa Y., et al. (2016). Genome-wide characterization of major intrinsic proteins in four grass plants and their non-aqua transport selectivity profiles with comparative perspective. PLoS ONE 11:e0157735 10.1371/journal.pone.0157735 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous