

Narrow-Leafed Lupin (Lupinus angustifolius) β1- and β6-Conglutin Proteins Exhibit Antifungal Activity, Protecting Plants against Necrotrophic Pathogen Induced Damage from Sclerotinia sclerotiorum and Phytophthora nicotianae
- PMID: 28018392
- PMCID: PMC5161055
- DOI: 10.3389/fpls.2016.01856
Narrow-Leafed Lupin (Lupinus angustifolius) β1- and β6-Conglutin Proteins Exhibit Antifungal Activity, Protecting Plants against Necrotrophic Pathogen Induced Damage from Sclerotinia sclerotiorum and Phytophthora nicotianae
Abstract
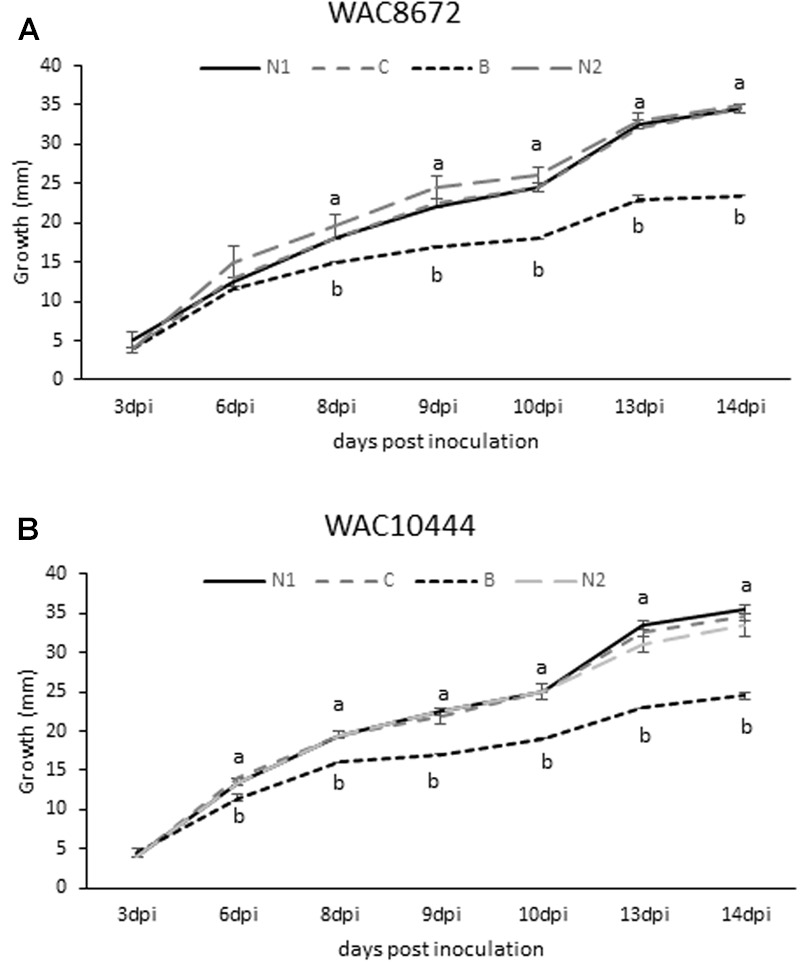
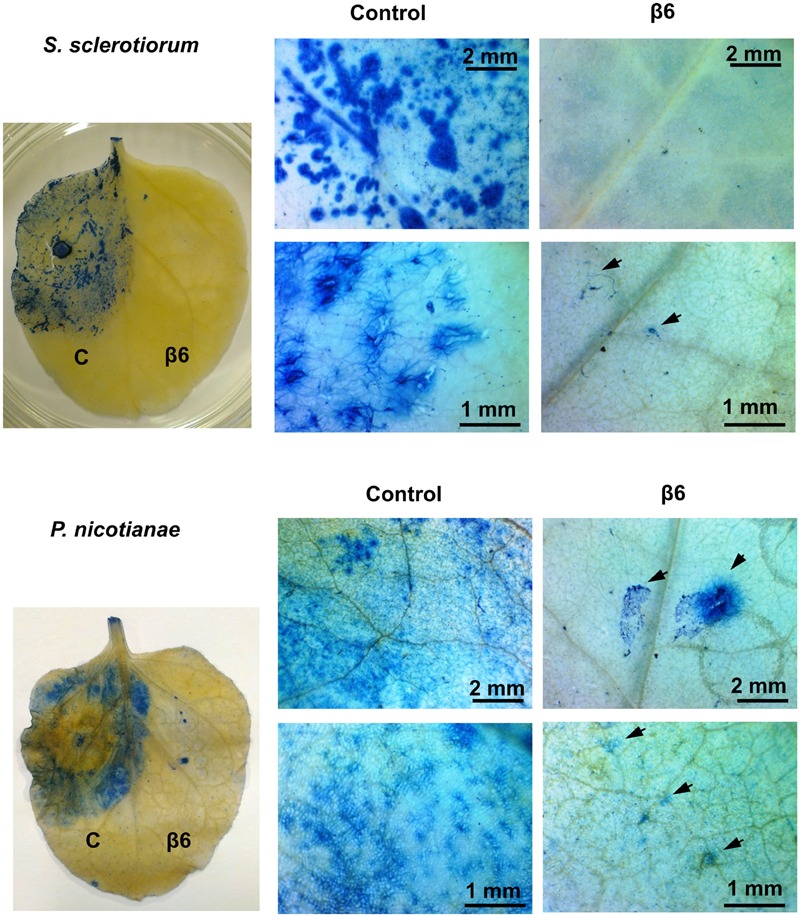
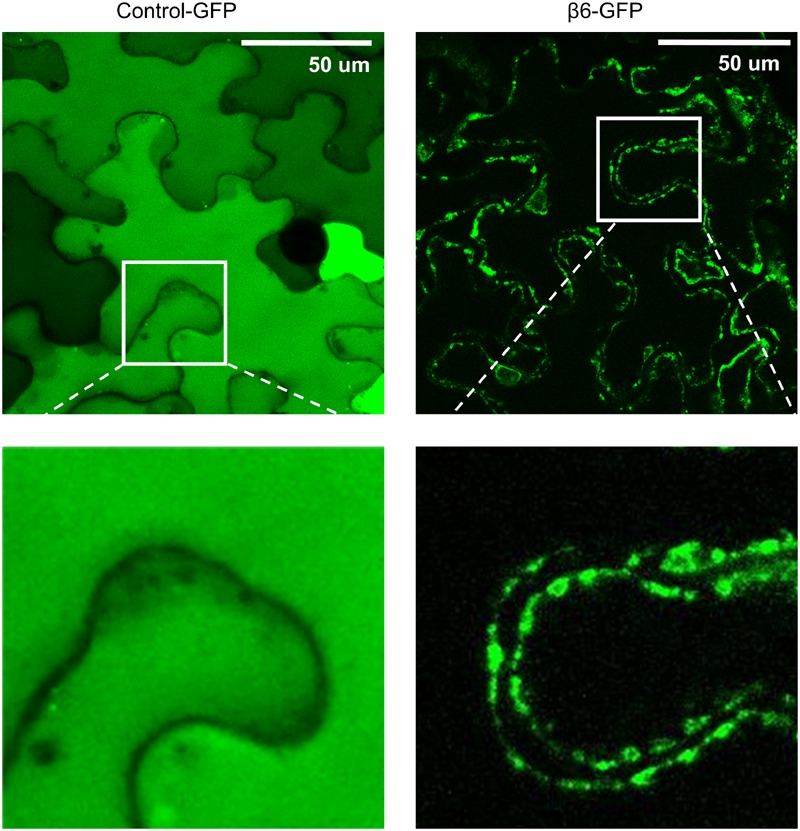
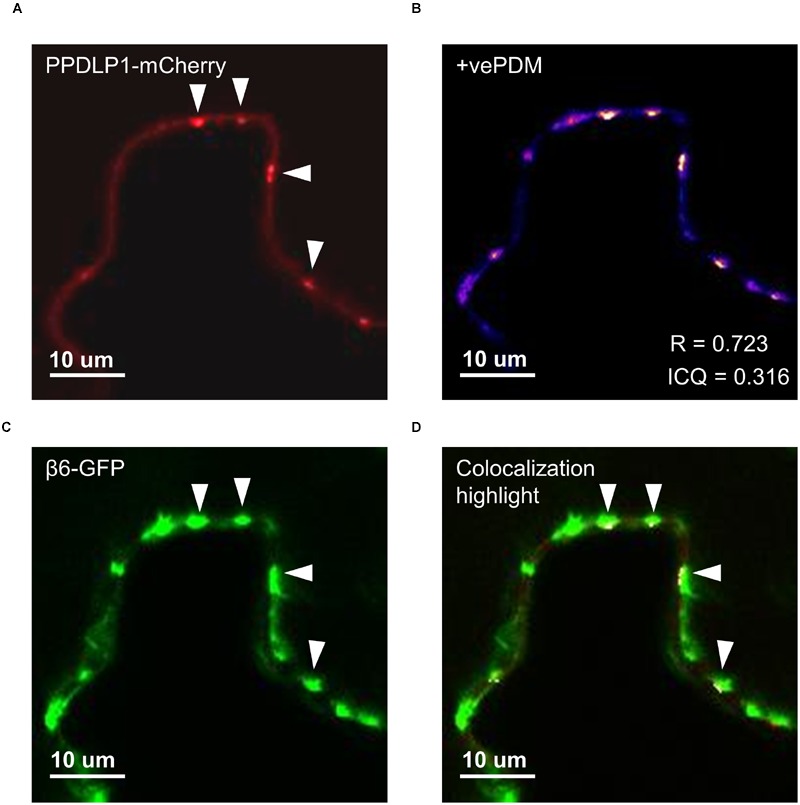
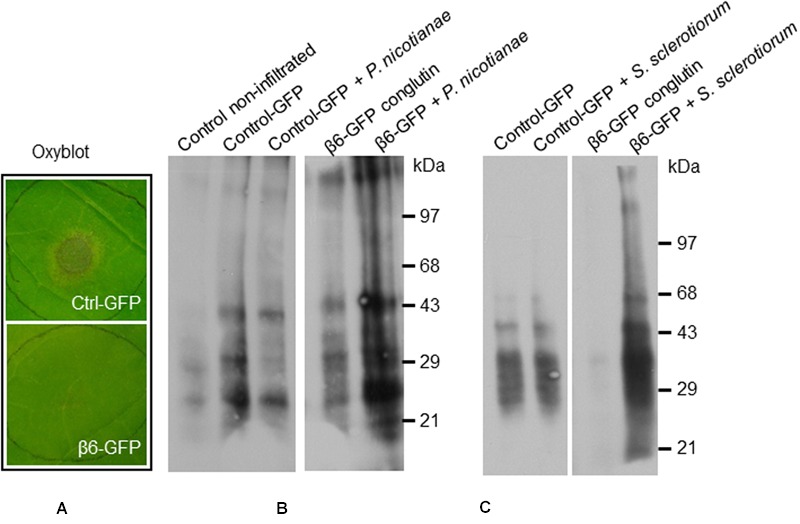
Vicilins (7S globulins) are seed storage proteins and constitute the main protein family in legume seeds, particularly in narrow-leafed lupin (Lupinus angustifolius L.; NLL), where seven vicilin genes, called β1- to β7-conglutin have been identified. Vicilins are involved in germination processes supplying amino acids for seedling growth and plant development, as well as in some cases roles in plant defense and protection against pathogens. The roles of NLL β-conglutins in plant defense are unknown. Here the potential role of five NLL β-conglutin family members in protection against necrotrophic fungal pathogens was investigated and it was demonstrated that recombinant purified 6xHis-tagged β1- and β6-conglutin proteins exhibited the strongest in vitro growth inhibitory activity against a range of necrotrophic fungal pathogens compared to β2, β3, and β4 conglutins. To examine activity in vivo, two representative necrotrophic pathogens, the fungus Sclerotinia sclerotiorum and oomycete Phytophthora nicotianae were used. Transient expression of β1- and β6-conglutin proteins in Nicotiana benthamiana leaves demonstrated in vivo growth suppression of both of these pathogens, resulting in low percentages of hyphal growth and elongation in comparison to control treated leaves. Cellular studies using β1- and β6-GFP fusion proteins showed these conglutins localized to the cell surface including plasmodesmata. Analysis of cellular death following S. sclerotiorum or P. nicotianae revealed both β1- and β6-conglutins suppressed pathogen induced cell death in planta and prevented pathogen induced suppression of the plant oxidative burst as determined by protein oxidation in infected compared to mock-inoculated leaves.
Keywords: 7S globulins; fungal pathogen; legume; oxidative stress; plant defense; seed storage protein; vicilins.
Figures

References
-
- Agrios G. N. (2005). “Plant diseases caused by fungi,” in Plant Pathology 5th Edn ed. Agrios G. N. (San Diego, CA: Academic Press; ) 385–614.
-
- Alscher R. G., Donahue J. L., Cramer C. L. (1997). Reactive oxygen species and antioxidants: Relationships in green cells. Physiol. Plant. 100 224–233. 10.1111/j.1399-3054.1997.tb04778.x - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources