

Postnatal ontogeny and the evolution of macrostomy in snakes
- PMID: 28018652
- PMCID: PMC5180150
- DOI: 10.1098/rsos.160612
Postnatal ontogeny and the evolution of macrostomy in snakes
Abstract
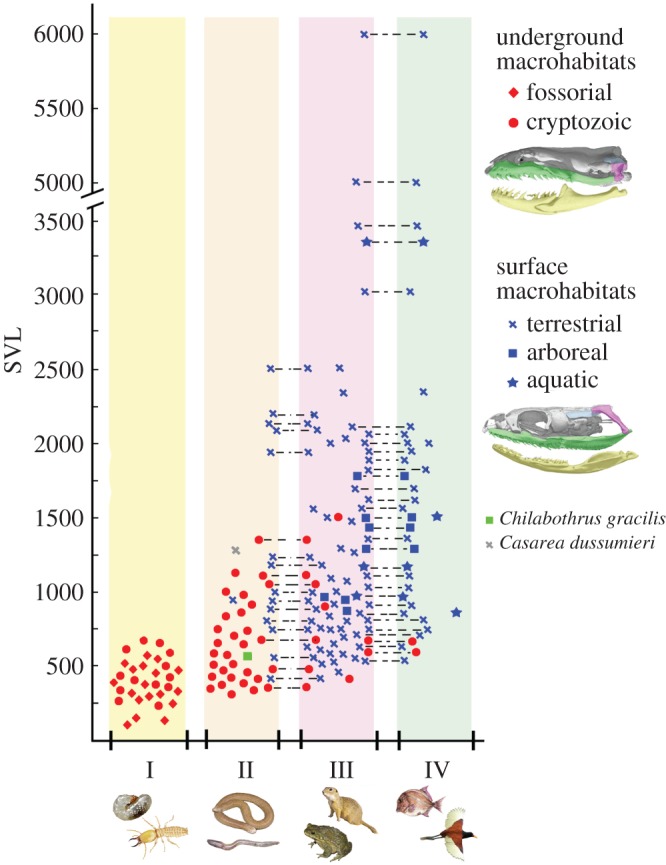
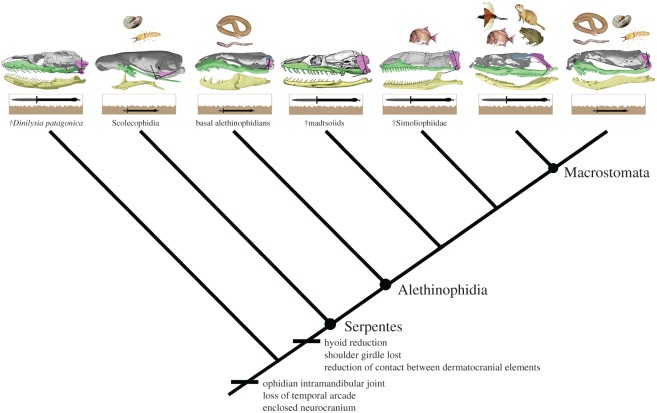
Macrostomy is the anatomical feature present in macrostomatan snakes that permits the ingestion of entire prey with high cross-sectional area. It depends on several anatomical traits in the skeleton and soft tissues, of which the elongation of gnathic complex and backward rotation of the quadrate represent crucial skeletal requirements. Here, the relevance of postnatal development of these skull structures and their relationship with macrohabitat and diet are explored. Contrary to the condition present in lizards and basal snakes that occupy underground macrohabitats, elements of the gnathic complex of most macrostomatan snakes that exploit surface macrohabitats display conspicuous elongation during postnatal growth, relative to the rest of the skull, as well as further backward rotation of the quadrate bone. Remarkably, several clades of small cryptozoic macrostomatans reverse these postnatal transformations and return to a diet based on prey with low cross-sectional area such as annelids, insects or elongated vertebrates, thus resembling the condition present in underground basal snakes. Dietary ontogenetic shift observed in most macrostomatan snakes is directly linked with this ontogenetic trajectory, indicating that this shift is acquired progressively as the gnathic complex elongates and the quadrate rotates backward during postnatal ontogeny. The numerous independent events of reversion in the gnathic complex and prey type choice observed in underground macrostomatans and the presence of skeletal requirements for macrostomy in extinct non-macrostomatan species reinforce the possibility that basal snakes represent underground survivors of clades that had the skeletal requirements for macrostomy. Taken together, the data presented here suggest that macrostomy has been shaped during multiple episodes of occupation of underground and surface macrohabitats throughout the evolution of snakes.
Keywords: evolution; macrostomy; postnatal ontogeny; snakes.
Figures
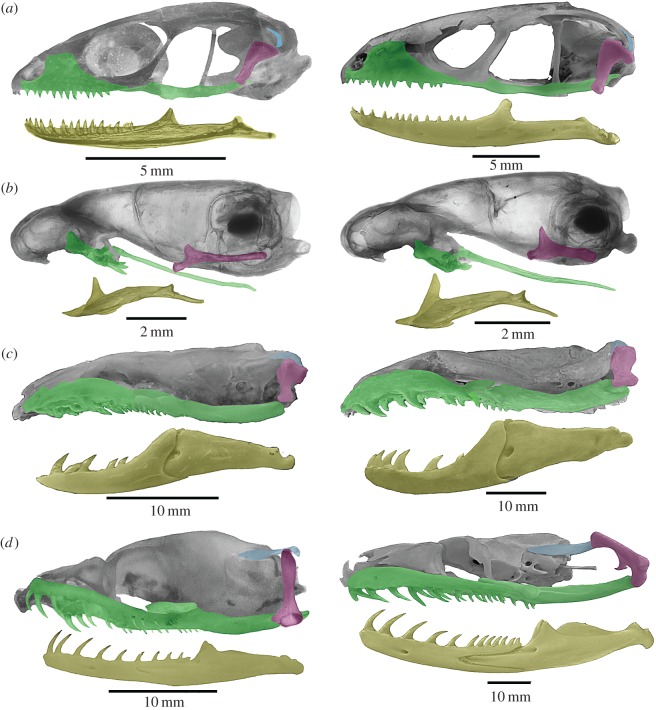
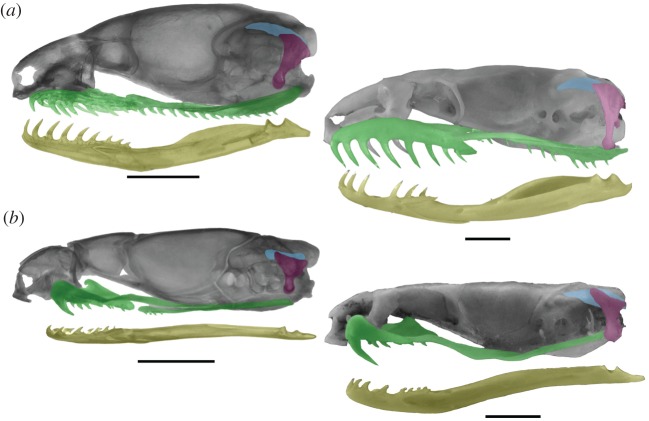
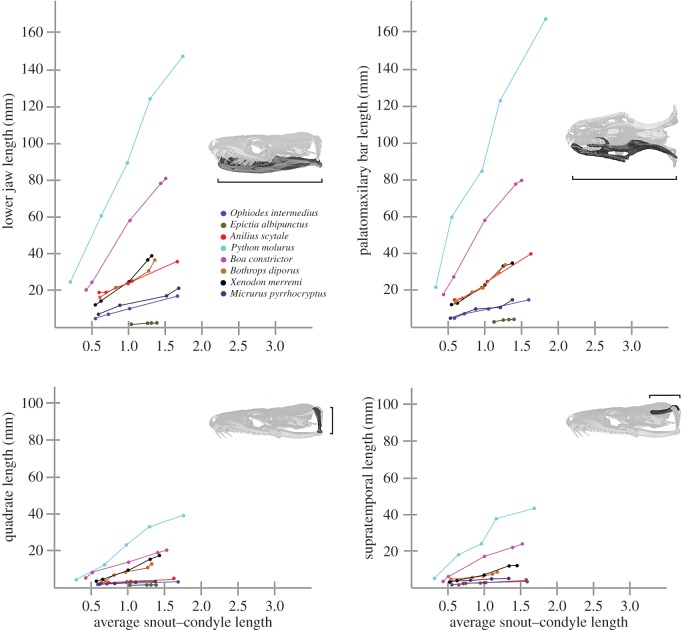
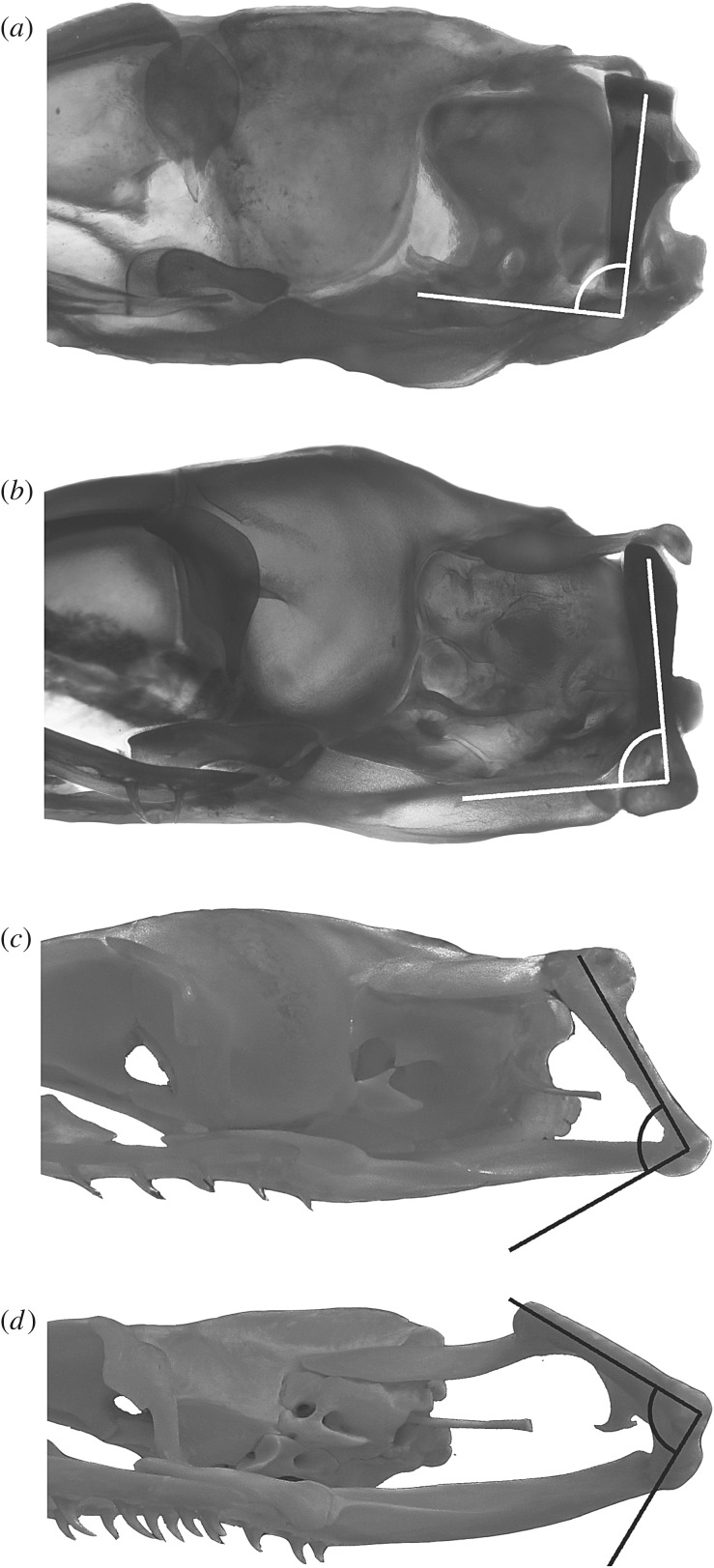
References
-
- Cundall D, Greene HW. 2000. Feeding in snakes. In Feeding: form, function and evolution in tetrapod vertebrates (ed. Schwenk K.), pp. 293–333. San Diego, CA: Academic Press.
-
- Cundall DF, Irish F. 2008. The snake skull. In Biology of the reptilia, Vol. 20, morphology H, the skull of Lepidosauria (eds Gans C, Gaunt AS, Adler K), pp. 349–692. Ithaca, NY: Society for the Study of Amphibians and Reptiles.
-
- Gans C. 1961. The feeding mechanism of snakes and its possible evolution. Am. Zool. 1, 217–227. (doi:10.1093/icb/1.2.217217-227) - DOI
-
- Rieppel O. 1980. The evolution of the ophidian feeding system. Zool. J. (Anat.) 103, 551–564.
-
- Young BA. 1998. The comparative morphology of the intermandibular connective tisssue in snakes (Reptilia: Squamata). Zool. Anz. 237, 59–84.
LinkOut - more resources
Full Text Sources
Other Literature Sources