

The TCF1-Bcl6 axis counteracts type I interferon to repress exhaustion and maintain T cell stemness
- PMID: 28018990
- PMCID: PMC5179228
- DOI: 10.1126/sciimmunol.aai8593
The TCF1-Bcl6 axis counteracts type I interferon to repress exhaustion and maintain T cell stemness
Abstract
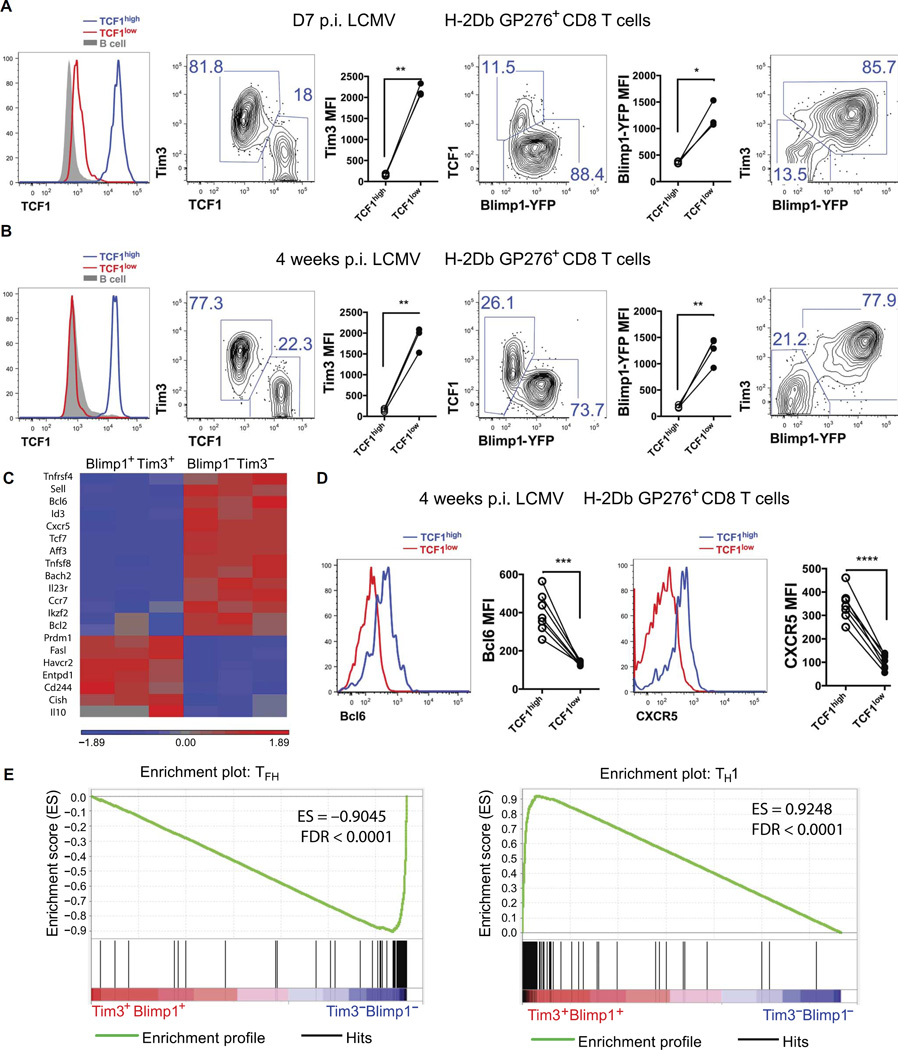
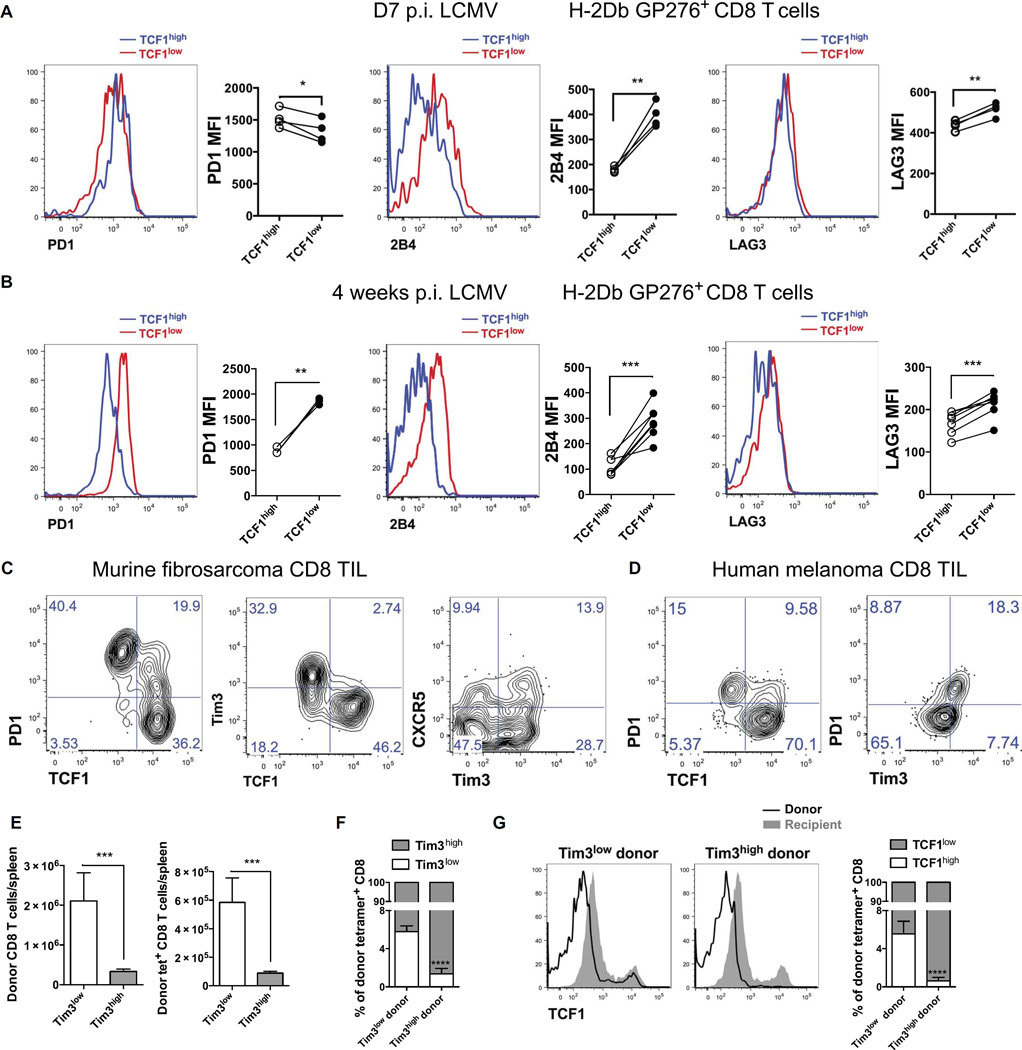
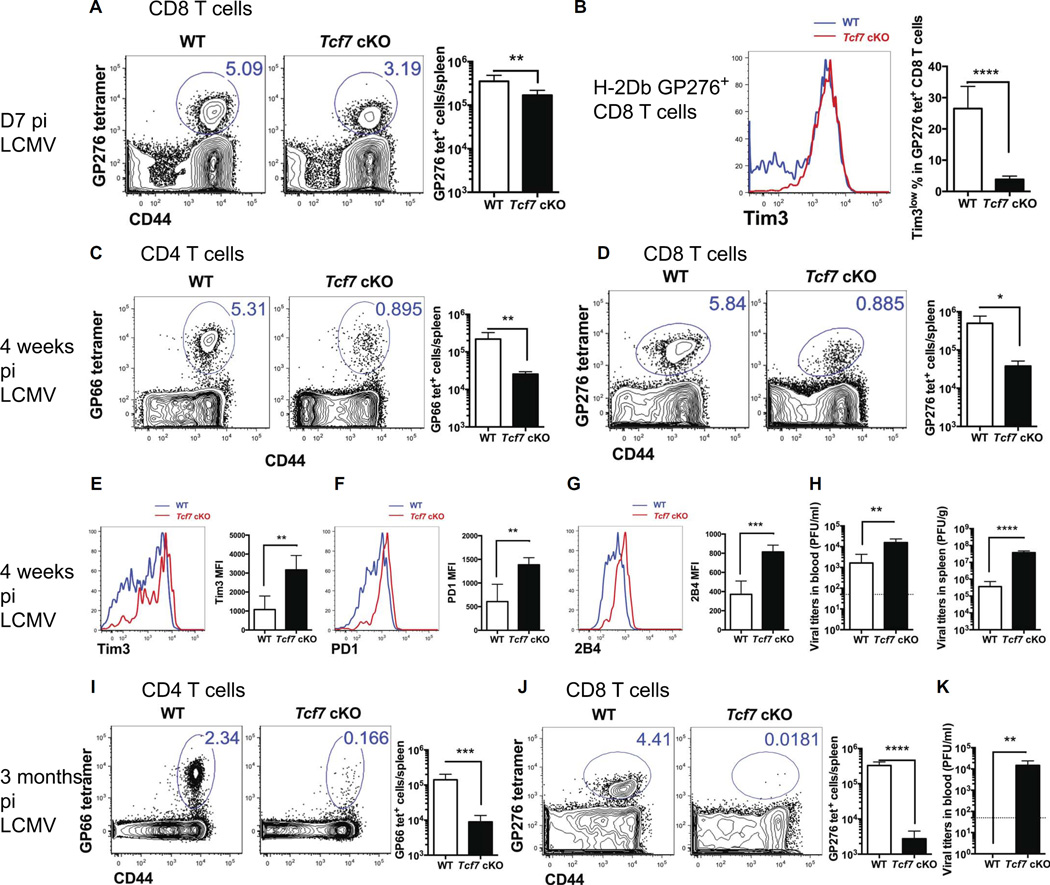
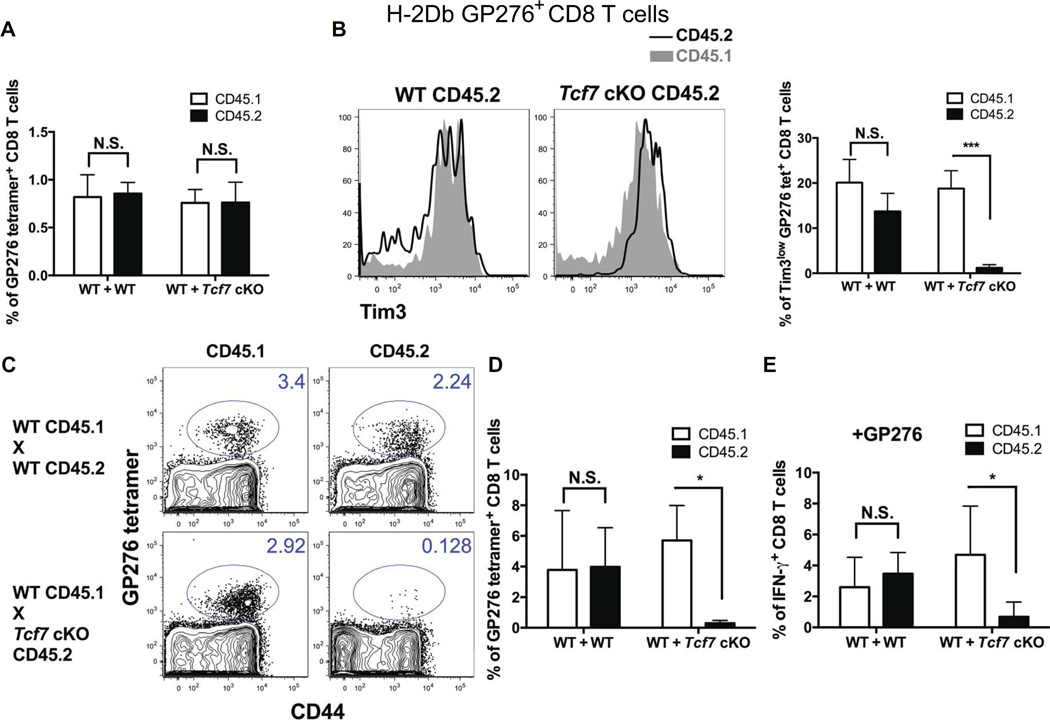
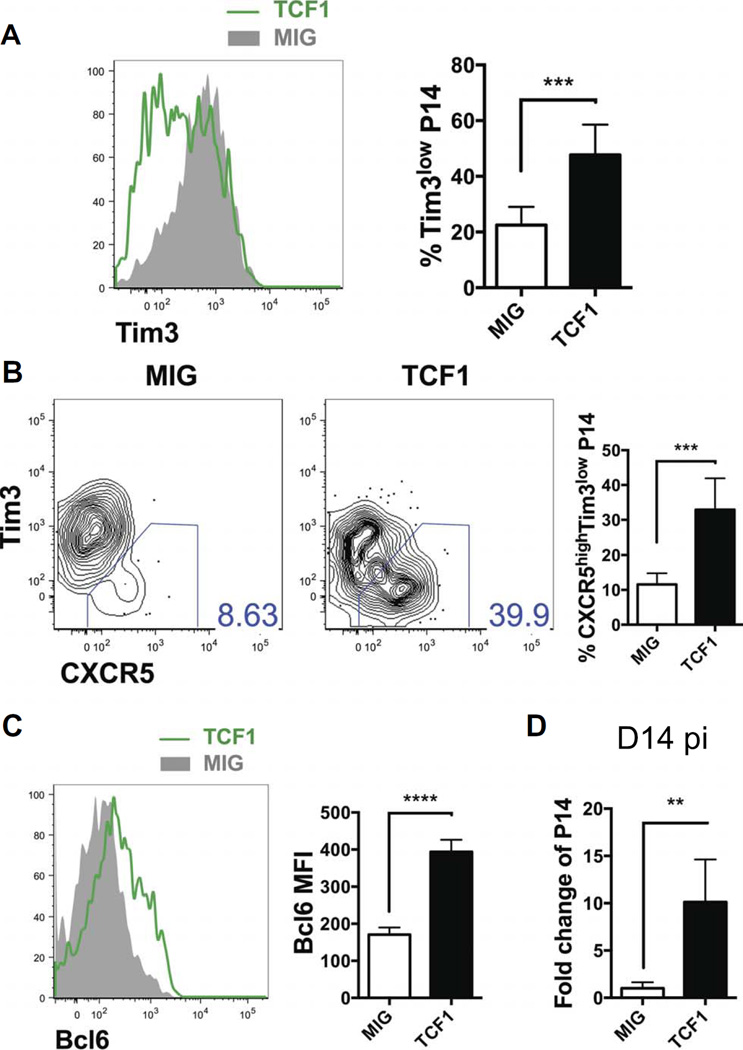
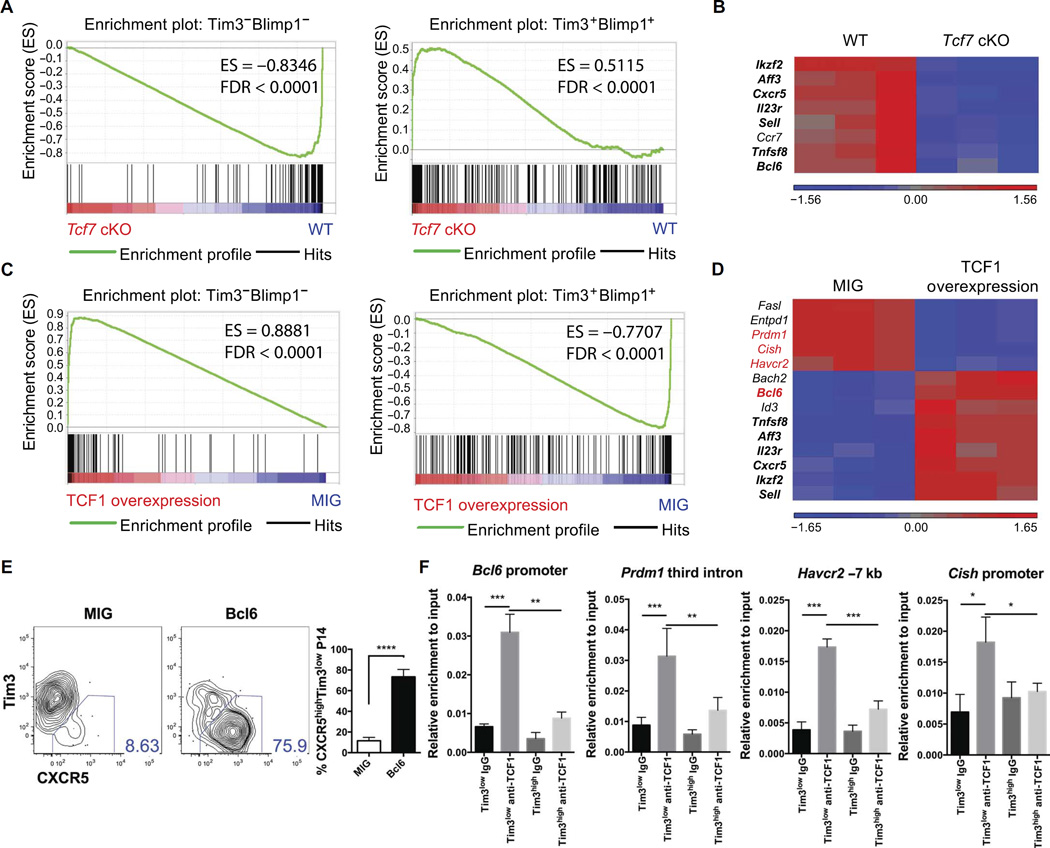
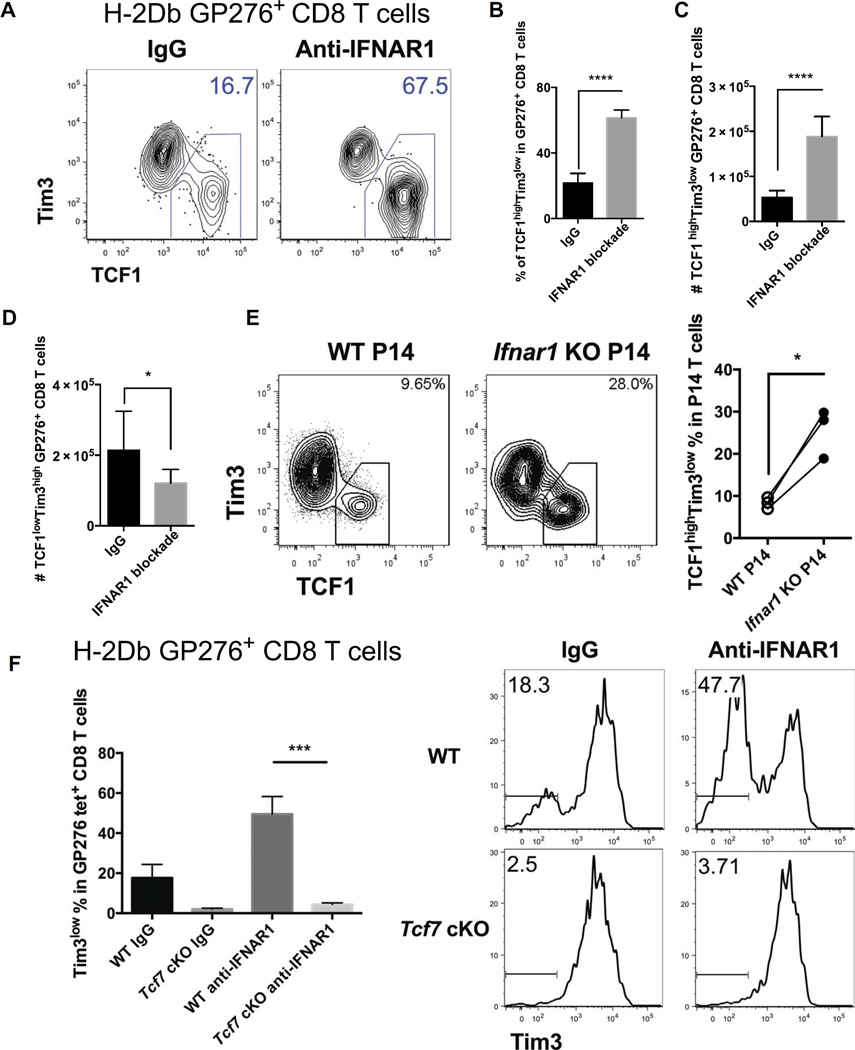
During chronic viral infections and in cancer, T cells become dysfunctional, a state known as T cell exhaustion. Although it is well recognized that memory CD8 T cells account for the persistence of CD8 T cell immunity after acute infection, how exhausted T cells persist remains less clear. Using chronic infection with lymphocytic choriomeningitis virus clone 13 and tumor samples, we demonstrate that CD8 T cells differentiate into a less exhausted TCF1high and a more exhausted TCF1low population. Virus-specific TCF1high CD8 T cells, which resemble T follicular helper (TFH) cells, persist and recall better than do TCF1low cells and act as progenitor cells to replenish TCF1low cells. We show that TCF1 is both necessary and sufficient to support this progenitor-like CD8 subset, whereas cell-intrinsic type I interferon signaling suppresses their differentiation. Accordingly, cell-intrinsic TCF1 deficiency led to a loss of these progenitor CD8 T cells, sharp contraction of virus-specific T cells, and uncontrolled viremia. Mechanistically, TCF1 repressed several pro-exhaustion factors and induced Bcl6 in CD8 T cells, which promoted the progenitor fate. We propose that the TCF1-Bcl6 axis counteracts type I interferon to repress T cell exhaustion and maintain T cell stemness, which is critical for persistent antiviral CD8 T cell responses in chronic infection. These findings provide insight into the requirements for persistence of T cell immune responses in the face of exhaustion and suggest mechanisms by which effective T cell-mediated immunity may be enhanced during chronic infections and cancer.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
