

Mutations in BOREALIN cause thyroid dysgenesis
- PMID: 28025328
- PMCID: PMC6311960
- DOI: 10.1093/hmg/ddw419
Mutations in BOREALIN cause thyroid dysgenesis
Abstract
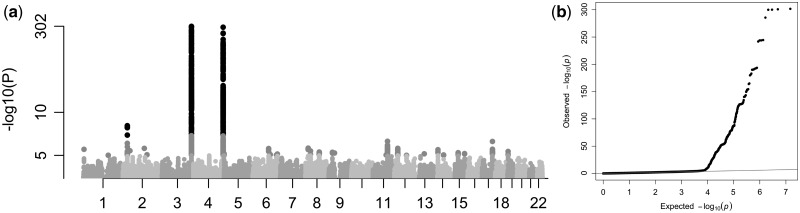
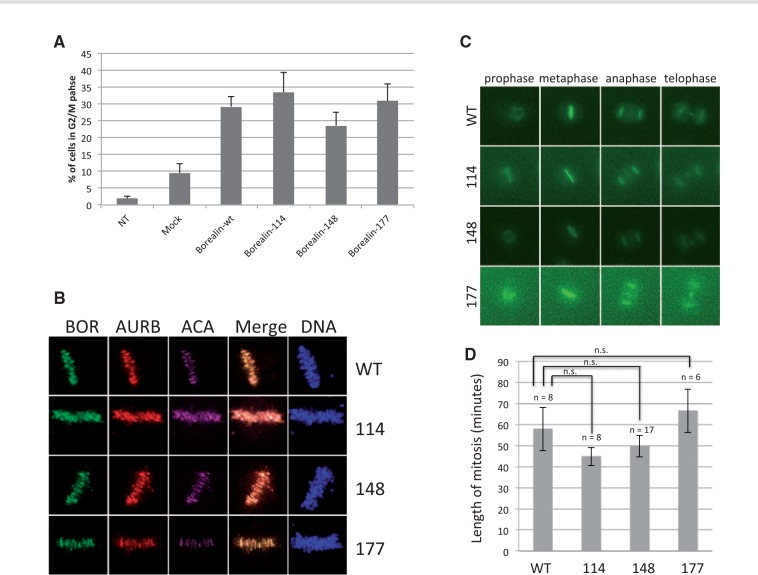
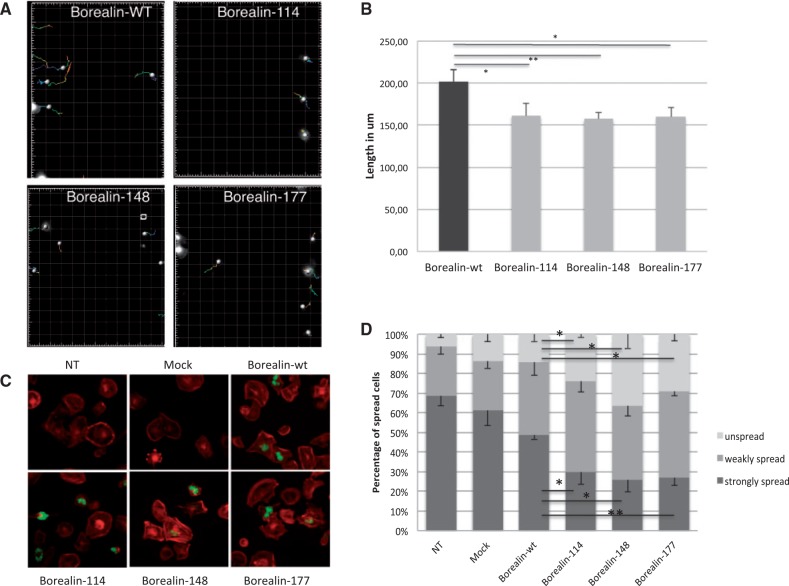
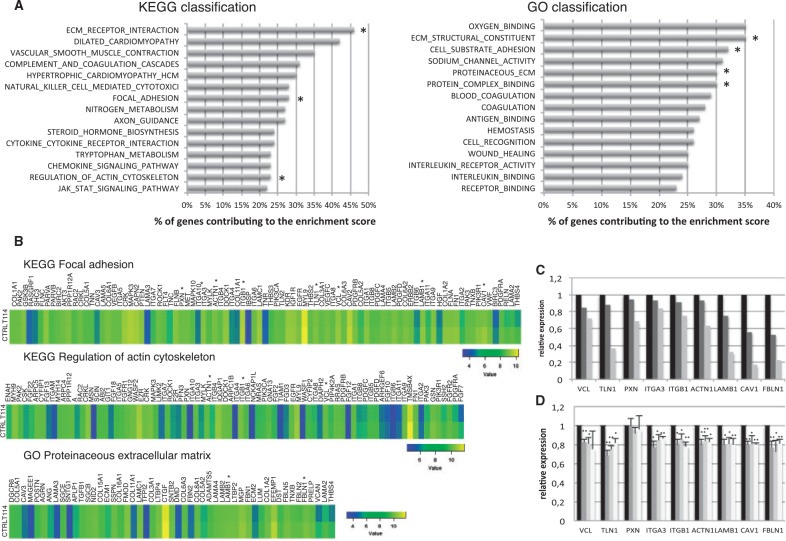
Congenital hypothyroidism is the most common neonatal endocrine disorder and is primarily caused by developmental abnormalities otherwise known as thyroid dysgenesis (TD). We performed whole exome sequencing (WES) in a consanguineous family with TD and subsequently sequenced a cohort of 134 probands with TD to identify genetic factors predisposing to the disease. We identified the novel missense mutations p.S148F, p.R114Q and p.L177W in the BOREALIN gene in TD-affected families. Borealin is a major component of the Chromosomal Passenger Complex (CPC) with well-known functions in mitosis. Further analysis of the missense mutations showed no apparent effects on mitosis. In contrast, expression of the mutants in human thyrocytes resulted in defects in adhesion and migration with corresponding changes in gene expression suggesting others functions for this mitotic protein. These results were well correlated with the same gene expression pattern analysed in the thyroid tissue of the patient with BOREALIN-p.R114W. These studies open new avenues in the genetics of TD in humans.
Trial registration: ClinicalTrials.gov NCT01916018.
© The Authors 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Stoupa A., Kariyawasam D., Carré A., Polak M. (2016) Update of Thyroid Developmental Genes. Endocrinol. Metab. Clin. North. Am., 45, 243–254. - PubMed
-
- Léger J., Marinovic D., Garel C., Bonaıïti-Pellié C., Polak M., Czernichow P. (2002) Thyroid developmental anomalies in first degree relatives of children with congenital hypothyroidism. J. Clin. Endocrinol. Metab., 87, 575–580. - PubMed
-
- Damante G., Tell G., Di Lauro R. (2001) A unique combination of transcription factors controls differentiation of thyroid cells. Prog. Nucleic. Acid. Res. Mol. Biol., 66, 307–356. - PubMed
-
- Fagman H., Nilsson M. (2010) Morphogenesis of the thyroid gland. Mol. Cell. Endocrinol., 35–54. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
