

The Hamiltonian Brain: Efficient Probabilistic Inference with Excitatory-Inhibitory Neural Circuit Dynamics
- PMID: 28027294
- PMCID: PMC5189947
- DOI: 10.1371/journal.pcbi.1005186
The Hamiltonian Brain: Efficient Probabilistic Inference with Excitatory-Inhibitory Neural Circuit Dynamics
Abstract
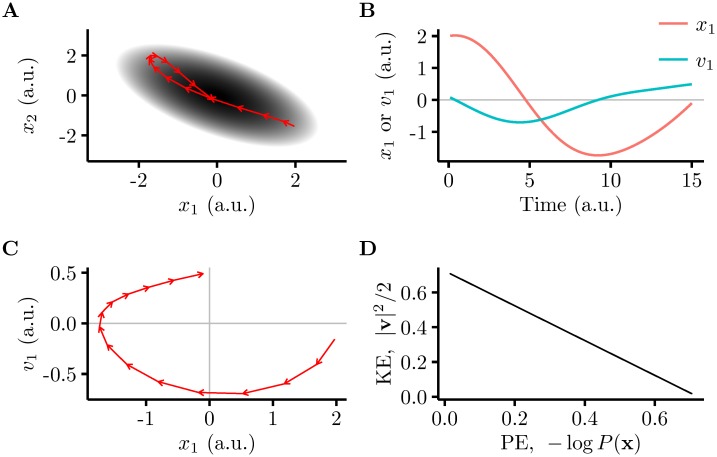
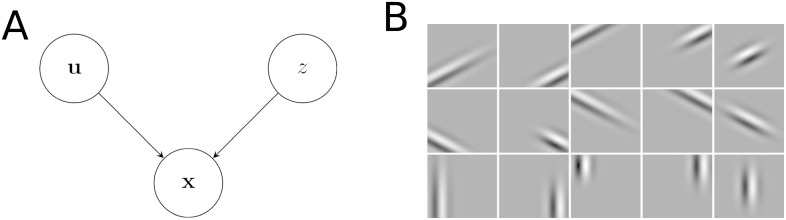
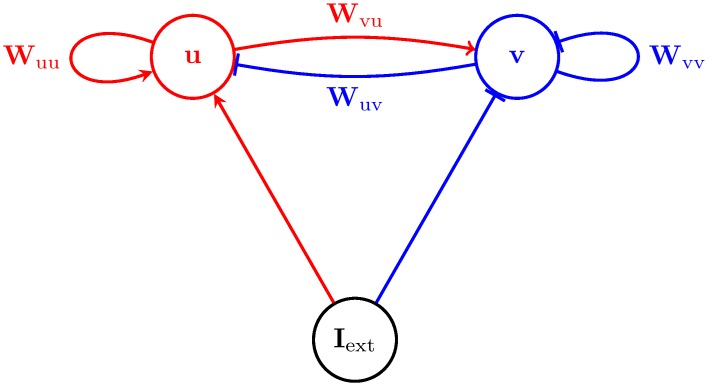
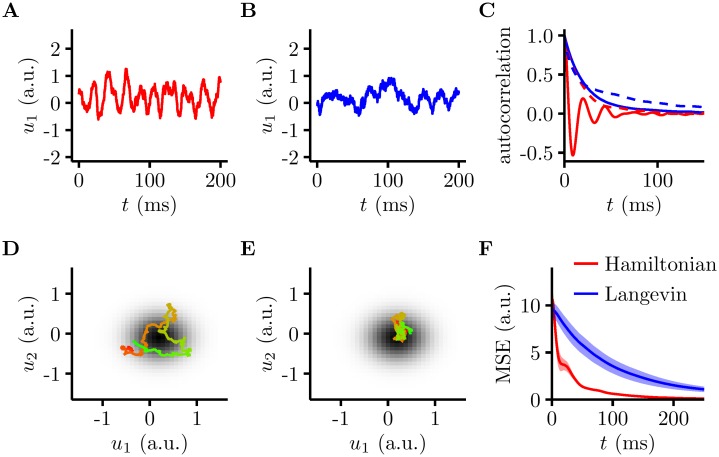
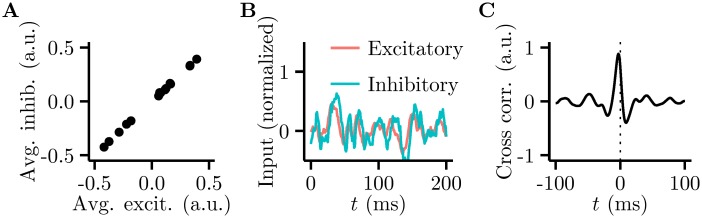
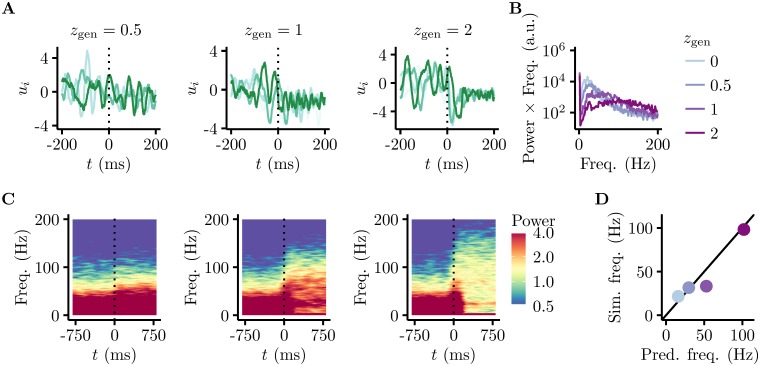
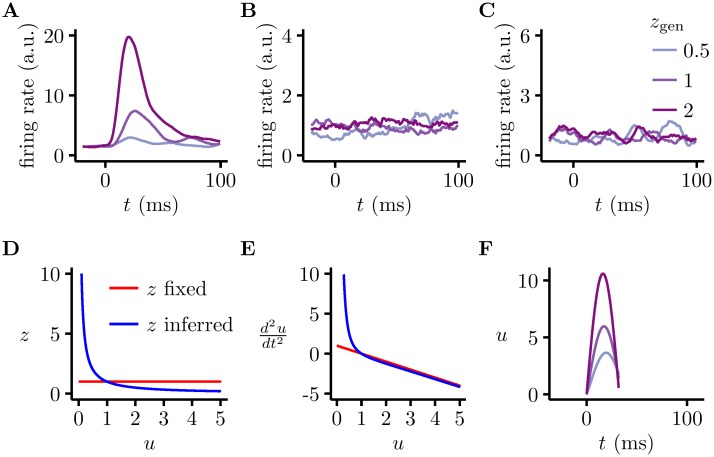
Probabilistic inference offers a principled framework for understanding both behaviour and cortical computation. However, two basic and ubiquitous properties of cortical responses seem difficult to reconcile with probabilistic inference: neural activity displays prominent oscillations in response to constant input, and large transient changes in response to stimulus onset. Indeed, cortical models of probabilistic inference have typically either concentrated on tuning curve or receptive field properties and remained agnostic as to the underlying circuit dynamics, or had simplistic dynamics that gave neither oscillations nor transients. Here we show that these dynamical behaviours may in fact be understood as hallmarks of the specific representation and algorithm that the cortex employs to perform probabilistic inference. We demonstrate that a particular family of probabilistic inference algorithms, Hamiltonian Monte Carlo (HMC), naturally maps onto the dynamics of excitatory-inhibitory neural networks. Specifically, we constructed a model of an excitatory-inhibitory circuit in primary visual cortex that performed HMC inference, and thus inherently gave rise to oscillations and transients. These oscillations were not mere epiphenomena but served an important functional role: speeding up inference by rapidly spanning a large volume of state space. Inference thus became an order of magnitude more efficient than in a non-oscillatory variant of the model. In addition, the network matched two specific properties of observed neural dynamics that would otherwise be difficult to account for using probabilistic inference. First, the frequency of oscillations as well as the magnitude of transients increased with the contrast of the image stimulus. Second, excitation and inhibition were balanced, and inhibition lagged excitation. These results suggest a new functional role for the separation of cortical populations into excitatory and inhibitory neurons, and for the neural oscillations that emerge in such excitatory-inhibitory networks: enhancing the efficiency of cortical computations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- van Beers RJ, Sittig AC, van der Gon JJD. Integration of proprioceptive and visual position-information: An experimentally supported model. Journal of Neurophysiology. 1999;81:1355–1364. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
