

Model-based analysis of DNA replication profiles: predicting replication fork velocity and initiation rate by profiling free-cycling cells
- PMID: 28028072
- PMCID: PMC5287236
- DOI: 10.1101/gr.205849.116
Model-based analysis of DNA replication profiles: predicting replication fork velocity and initiation rate by profiling free-cycling cells
Abstract
Eukaryotic cells initiate DNA synthesis by sequential firing of hundreds of origins. This ordered replication is described by replication profiles, which measure the DNA content within a cell population. Here, we show that replication dynamics can be deduced from replication profiles of free-cycling cells. While such profiles lack explicit temporal information, they are sensitive to fork velocity and initiation capacity through the passive replication pattern, namely the replication of origins by forks emanating elsewhere. We apply our model-based approach to a compendium of profiles that include most viable budding yeast mutants implicated in replication. Predicted changes in fork velocity or initiation capacity are verified by profiling synchronously replicating cells. Notably, most mutants implicated in late (or early) origin effects are explained by global modulation of fork velocity or initiation capacity. Our approach provides a rigorous framework for analyzing DNA replication profiles of free-cycling cells.
© 2017 Gispan et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
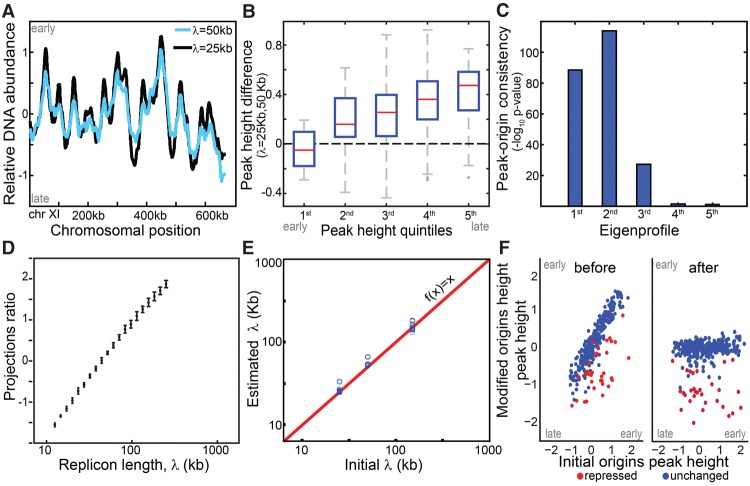
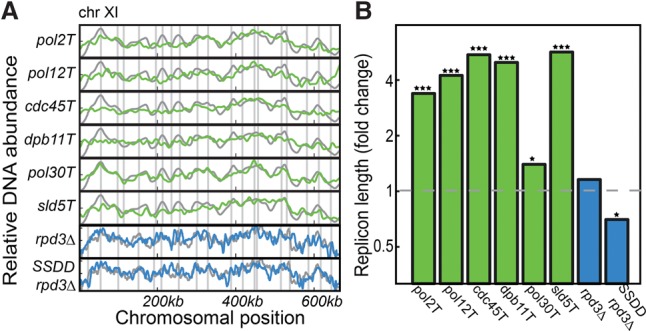
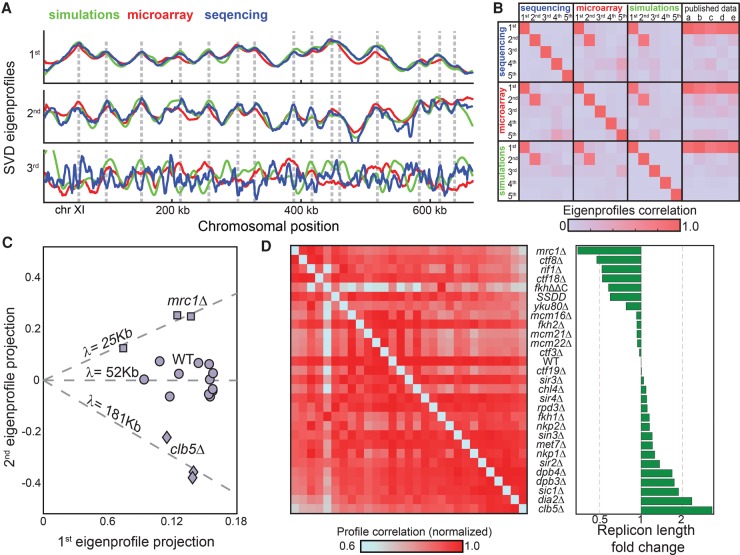
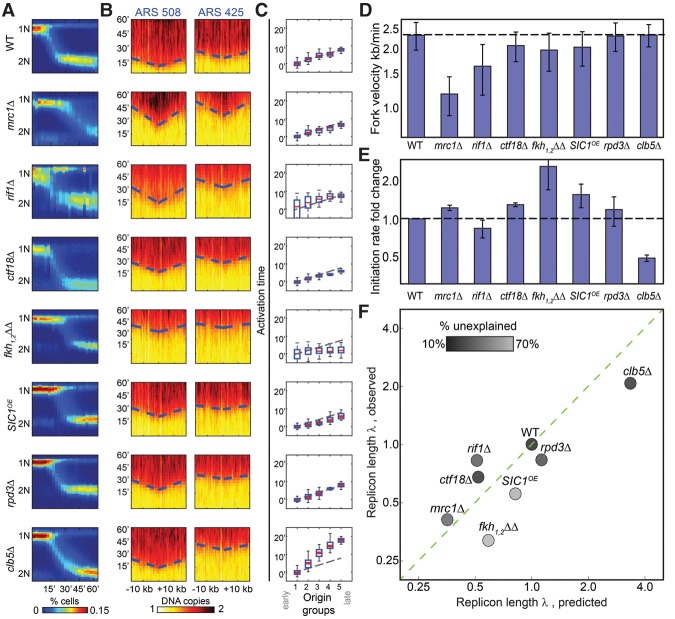
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases