

Adenosine Shifts Plasticity Regimes between Associative and Homeostatic by Modulating Heterosynaptic Changes
- PMID: 28028196
- PMCID: PMC5299565
- DOI: 10.1523/JNEUROSCI.2984-16.2016
Adenosine Shifts Plasticity Regimes between Associative and Homeostatic by Modulating Heterosynaptic Changes
Abstract
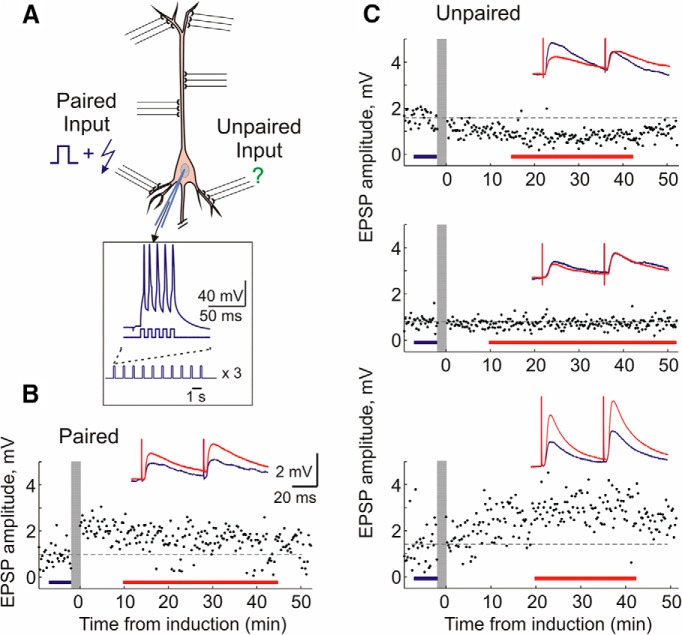
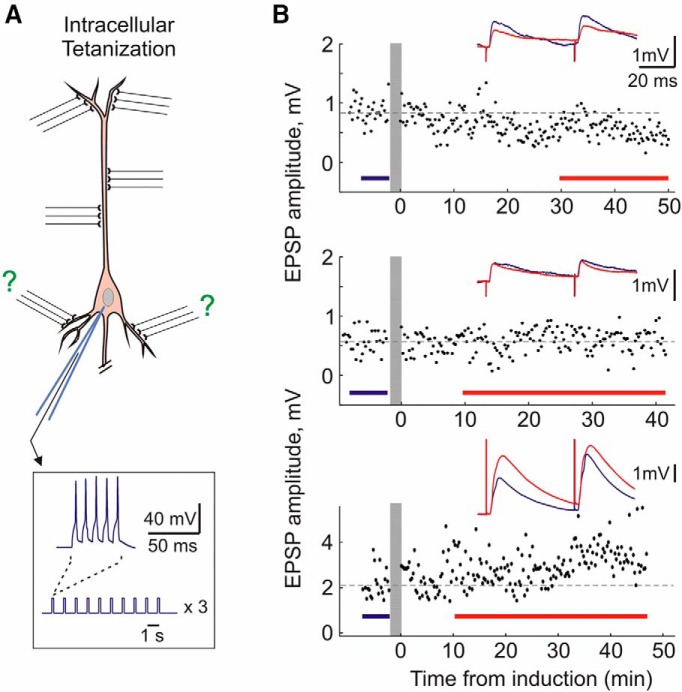
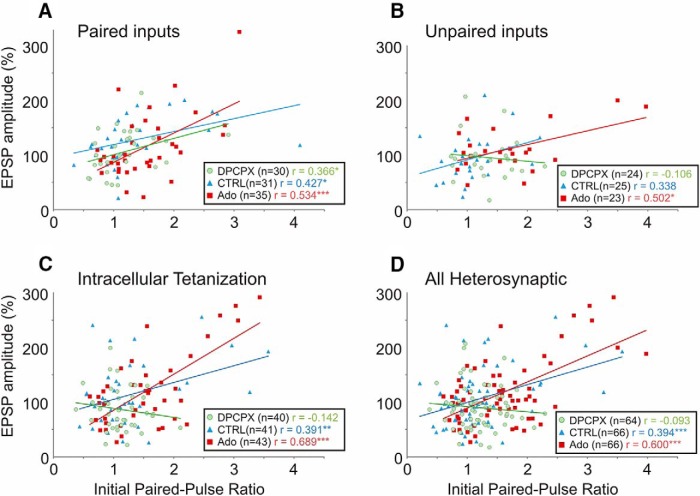
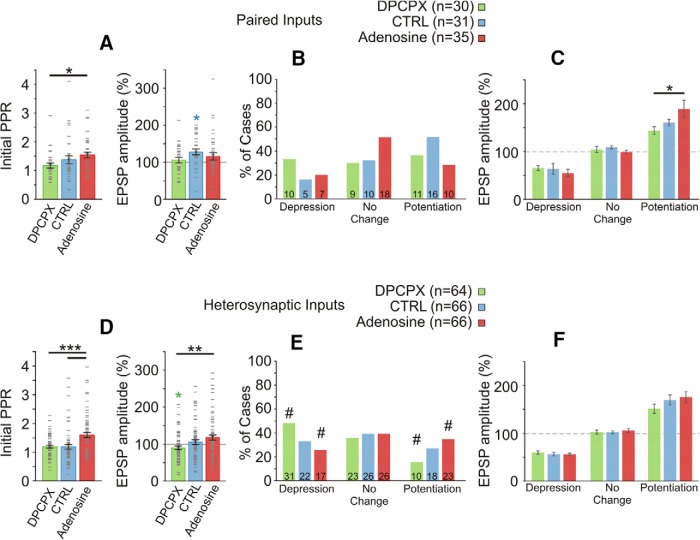
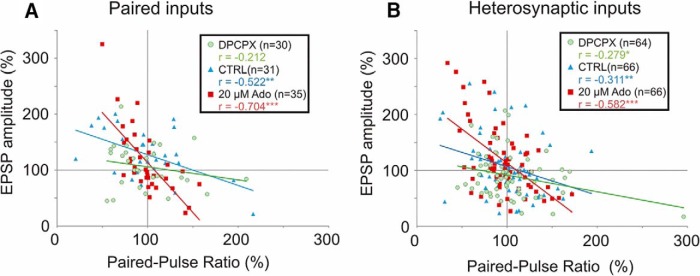
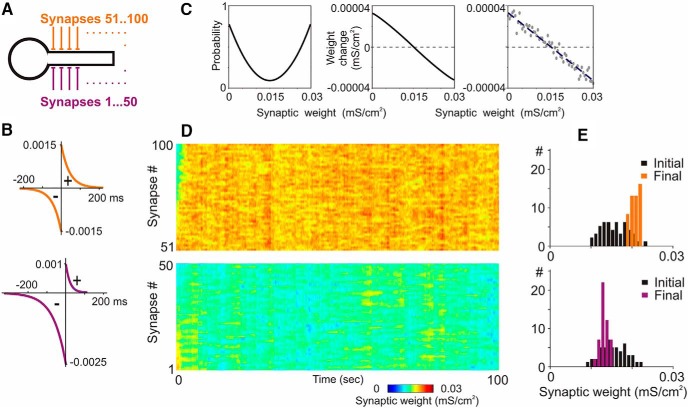
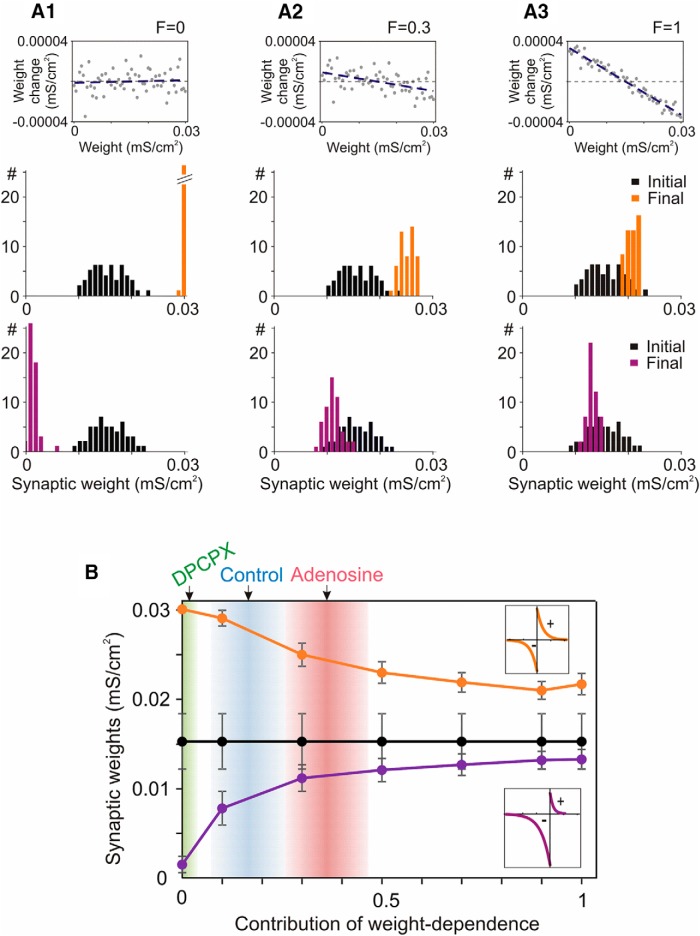
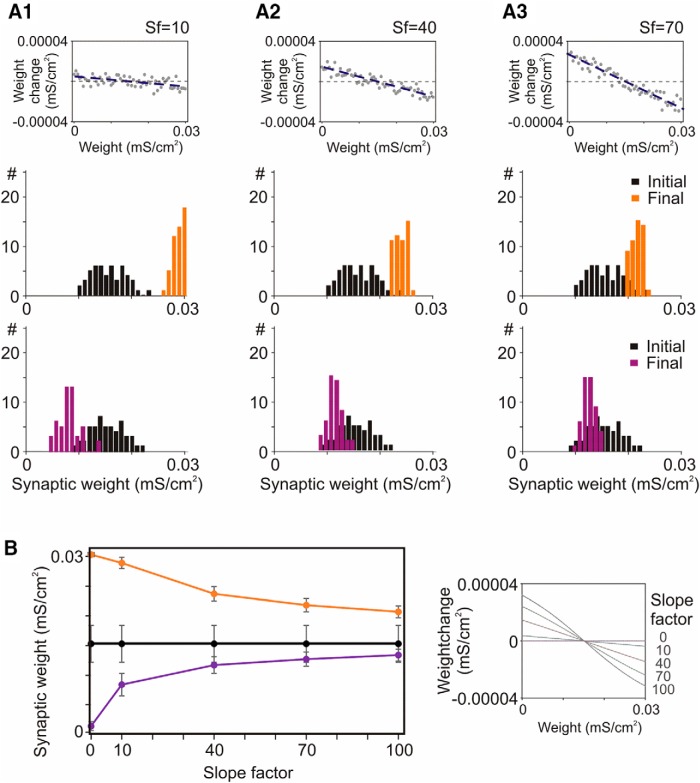
Endogenous extracellular adenosine level fluctuates in an activity-dependent manner and with sleep-wake cycle, modulating synaptic transmission and short-term plasticity. Hebbian-type long-term plasticity introduces intrinsic positive feedback on synaptic weight changes, making them prone to runaway dynamics. We previously demonstrated that co-occurring, weight-dependent heterosynaptic plasticity can robustly prevent runaway dynamics. Here we show that at neocortical synapses in slices from rat visual cortex, adenosine modulates the weight dependence of heterosynaptic plasticity: blockade of adenosine A1 receptors abolished weight dependence, while increased adenosine level strengthened it. Using model simulations, we found that the strength of weight dependence determines the ability of heterosynaptic plasticity to prevent runaway dynamics of synaptic weights imposed by Hebbian-type learning. Changing the weight dependence of heterosynaptic plasticity within an experimentally observed range gradually shifted the operating point of neurons between an unbalancing regime dominated by associative plasticity and a homeostatic regime of tightly constrained synaptic changes. Because adenosine tone is a natural correlate of activity level (activity increases adenosine tone) and brain state (elevated adenosine tone increases sleep pressure), modulation of heterosynaptic plasticity by adenosine represents an endogenous mechanism that translates changes of the brain state into a shift of the regime of synaptic plasticity and learning. We speculate that adenosine modulation may provide a mechanism for fine-tuning of plasticity and learning according to brain state and activity.SIGNIFICANCE STATEMENT Associative learning depends on brain state and is impaired when the subject is sleepy or tired. However, the link between changes of brain state and modulation of synaptic plasticity and learning remains elusive. Here we show that adenosine regulates weight dependence of heterosynaptic plasticity: adenosine strengthened weight dependence of heterosynaptic plasticity; blockade of adenosine A1 receptors abolished it. In model neurons, such changes of the weight dependence of heterosynaptic plasticity shifted their operating point between regimes dominated by associative plasticity or by synaptic homeostasis. Because adenosine tone is a natural correlate of activity level and brain state, modulation of plasticity by adenosine represents an endogenous mechanism for translation of brain state changes into a shift of the regime of synaptic plasticity and learning.
Keywords: adenosine; heterosynaptic plasticity; learning rules; neuron models; synaptic plasticity; visual cortex.
Copyright © 2017 the authors 0270-6474/17/371439-14$15.00/0.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources