

Barriers to Infection of Human Cells by Feline Leukemia Virus: Insights into Resistance to Zoonosis
- PMID: 28031367
- PMCID: PMC5309941
- DOI: 10.1128/JVI.02119-16
Barriers to Infection of Human Cells by Feline Leukemia Virus: Insights into Resistance to Zoonosis
Abstract
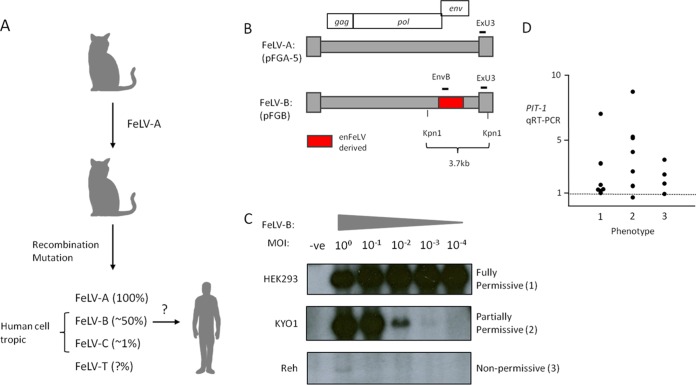
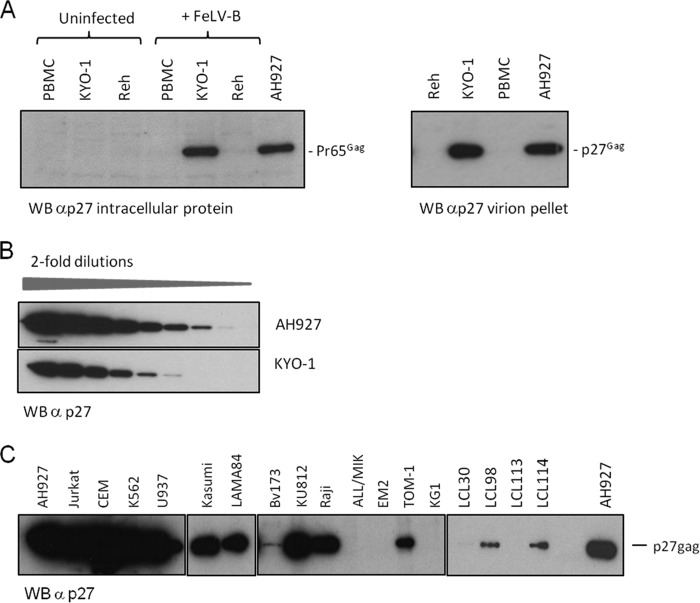
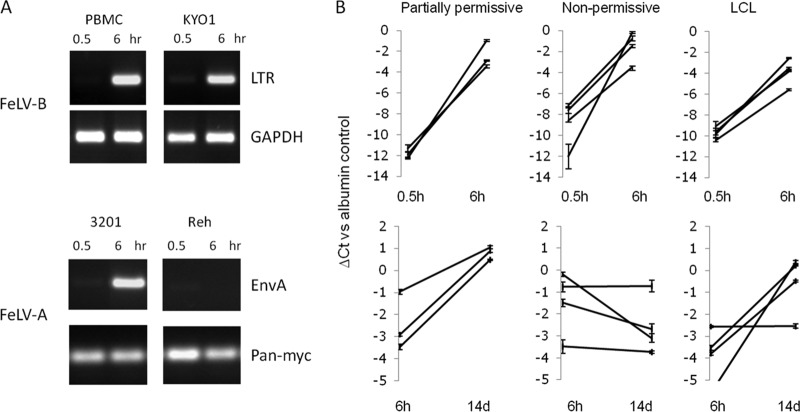
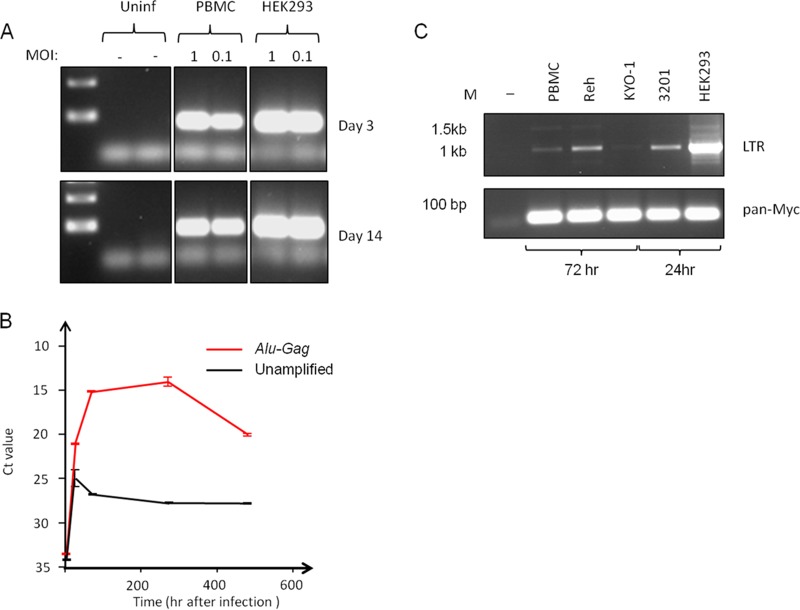
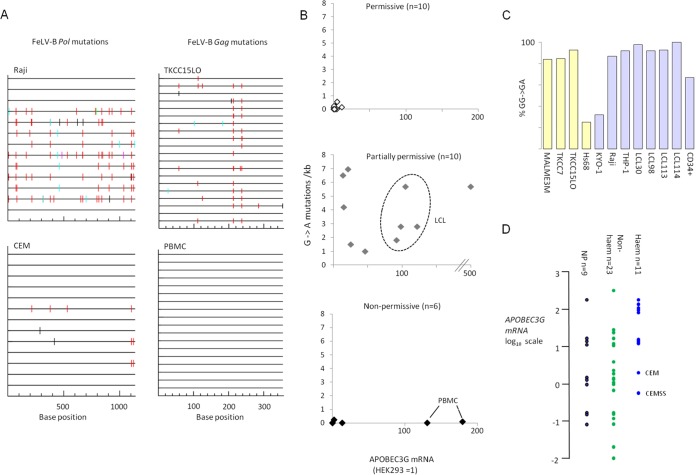
The human genome displays a rich fossil record of past gammaretrovirus infections, yet no current epidemic is evident, despite environmental exposure to viruses that infect human cells in vitro Feline leukemia viruses (FeLVs) rank high on this list, but neither domestic nor workplace exposure has been associated with detectable serological responses. Nonspecific inactivation of gammaretroviruses by serum factors appears insufficient to explain these observations. To investigate further, we explored the susceptibilities of primary and established human cell lines to FeLV-B, the most likely zoonotic variant. Fully permissive infection was common in cancer-derived cell lines but was also a feature of nontransformed keratinocytes and lung fibroblasts. Cells of hematopoietic origin were generally less permissive and formed discrete groups on the basis of high or low intracellular protein expression and virion release. Potent repression was observed in primary human blood mononuclear cells and a subset of leukemia cell lines. However, the early steps of reverse transcription and integration appear to be unimpaired in nonpermissive cells. FeLV-B was subject to G→A hypermutation with a predominant APOBEC3G signature in partially permissive cells but was not mutated in permissive cells or in nonpermissive cells that block secondary viral spread. Distinct cellular barriers that protect primary human blood cells are likely to be important in protection against zoonotic infection with FeLV.IMPORTANCE Domestic exposure to gammaretroviruses such as feline leukemia viruses (FeLVs) occurs worldwide, but the basis of human resistance to infection remains incompletely understood. The potential threat is evident from the human genome sequence, which reveals many past epidemics of gammaretrovirus infection, and from recent cross-species jumps of gammaretroviruses from rodents to primates and marsupials. This study examined resistance to infection at the cellular level with the most prevalent human cell-tropic FeLV variant, FeLV-B. We found that blood cells are uniquely resistant to infection with FeLV-B due to the activity of cellular enzymes that mutate the viral genome. A second block, which appears to suppress viral gene expression after the viral genome has integrated into the host cell genome, was identified. Since cells derived from other normal human cell types are fully supportive of FeLV replication, innate resistance of blood cells could be critical in protecting against cross-species infection.
Keywords: APOBEC; feline leukemia virus; restriction factors; zoonosis.
Copyright © 2017 Terry et al.
Figures
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J, Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, Sougnez C, Stange-Thomann Y, Stojanovic N, Subramanian A, Wyman D, Rogers J, Sulston J, Ainscough R, Beck S, Bentley D, Burton J, Clee C, Carter N, Coulson A, Deadman R, Deloukas P, Dunham A, Dunham I, Durbin R, French L, et al. ; International Human Genome Sequencing Consortium. 2001. Initial sequencing and analysis of the human genome. Nature 409:860–921. doi: 10.1038/35057062. - DOI - PubMed
-
- Olah Z, Lehel C, Anderson WB, Eiden MV, Wilson CA. 1994. The cellular receptor for gibbon ape leukemia virus is a novel high affinity sodium-dependent phosphate transporter. J Biol Chem 269:25426–25431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
