

Principles of auditory processing differ between sensory and premotor structures of the songbird forebrain
- PMID: 28031398
- PMCID: PMC5349330
- DOI: 10.1152/jn.00462.2016
Principles of auditory processing differ between sensory and premotor structures of the songbird forebrain
Abstract
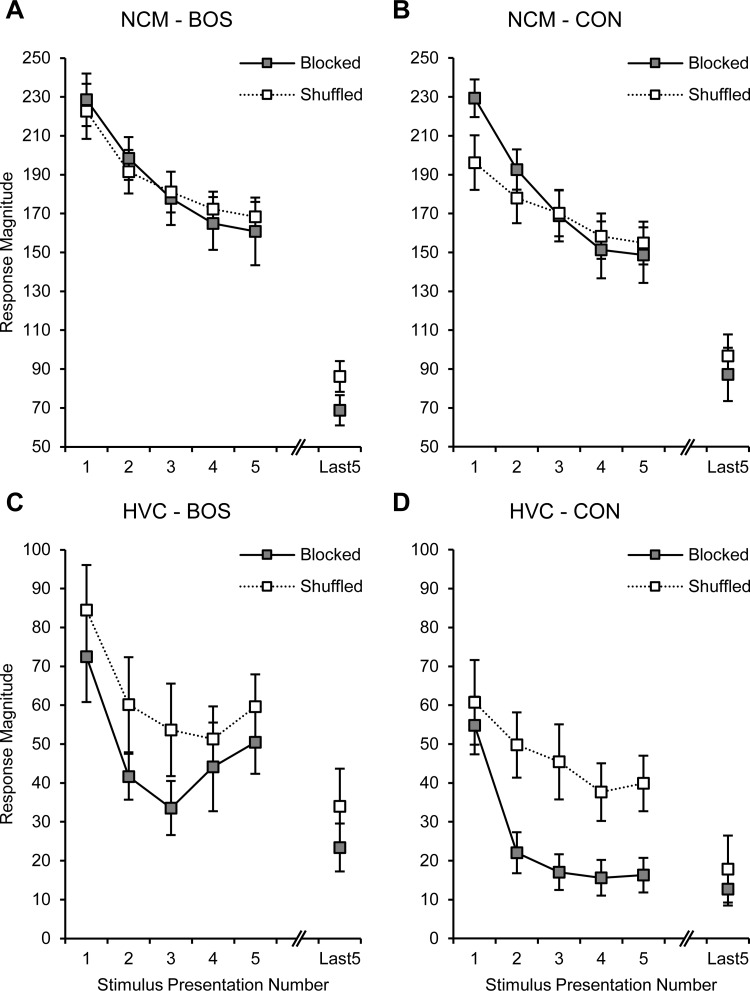
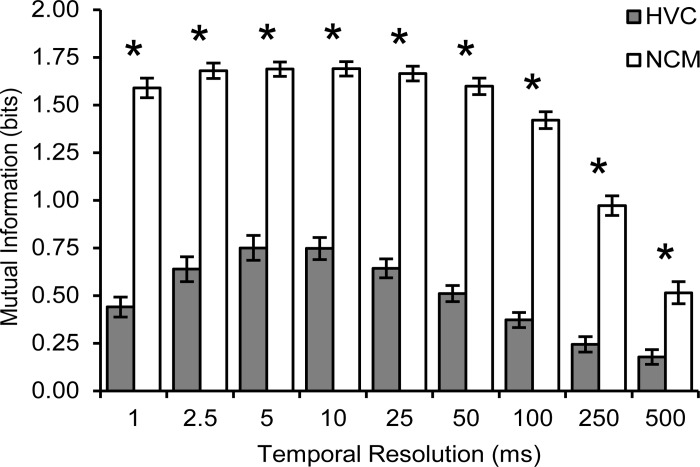
Sensory and motor brain structures work in collaboration during perception. To evaluate their respective contributions, the present study recorded neural responses to auditory stimulation at multiple sites simultaneously in both the higher-order auditory area NCM and the premotor area HVC of the songbird brain in awake zebra finches (Taeniopygia guttata). Bird's own song (BOS) and various conspecific songs (CON) were presented in both blocked and shuffled sequences. Neural responses showed plasticity in the form of stimulus-specific adaptation, with markedly different dynamics between the two structures. In NCM, the response decrease with repetition of each stimulus was gradual and long-lasting and did not differ between the stimuli or the stimulus presentation sequences. In contrast, HVC responses to CON stimuli decreased much more rapidly in the blocked than in the shuffled sequence. Furthermore, this decrease was more transient in HVC than in NCM, as shown by differential dynamics in the shuffled sequence. Responses to BOS in HVC decreased more gradually than to CON stimuli. The quality of neural representations, computed as the mutual information between stimuli and neural activity, was higher in NCM than in HVC. Conversely, internal functional correlations, estimated as the coherence between recording sites, were greater in HVC than in NCM. The cross-coherence between the two structures was weak and limited to low frequencies. These findings suggest that auditory communication signals are processed according to very different but complementary principles in NCM and HVC, a contrast that may inform study of the auditory and motor pathways for human speech processing.NEW & NOTEWORTHY Neural responses to auditory stimulation in sensory area NCM and premotor area HVC of the songbird forebrain show plasticity in the form of stimulus-specific adaptation with markedly different dynamics. These two structures also differ in stimulus representations and internal functional correlations. Accordingly, NCM seems to process the individually specific complex vocalizations of others based on prior familiarity, while HVC responses appear to be modulated by transitions and/or timing in the ongoing sequence of sounds.
Keywords: electrophysiology, songbird; mutual information; sensory-motor structures; stimulus-specific adaptation.
Copyright © 2017 the American Physiological Society.
Figures







References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources