

Force Spectroscopy of the Plasmodium falciparum Vaccine Candidate Circumsporozoite Protein Suggests a Mechanically Pliable Repeat Region
- PMID: 28031457
- PMCID: PMC5313086
- DOI: 10.1074/jbc.M116.754796
Force Spectroscopy of the Plasmodium falciparum Vaccine Candidate Circumsporozoite Protein Suggests a Mechanically Pliable Repeat Region
Abstract
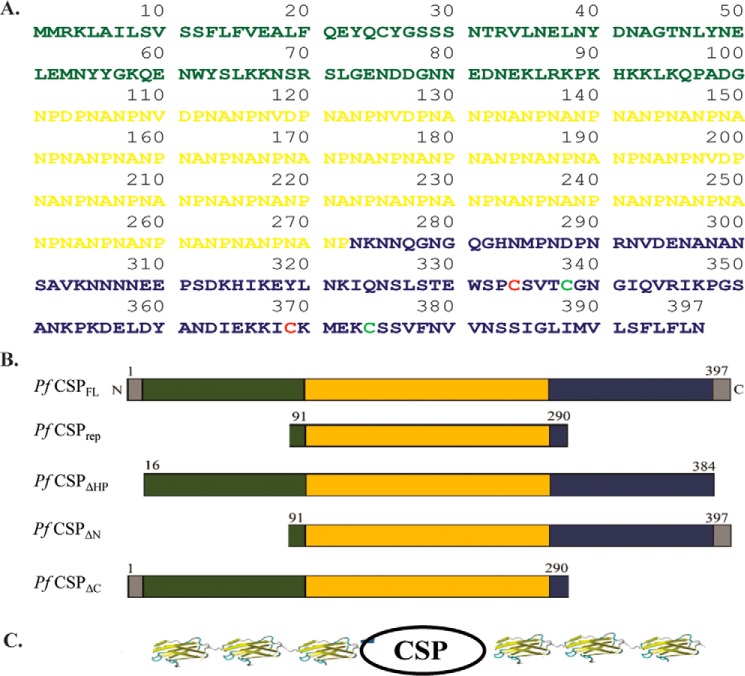
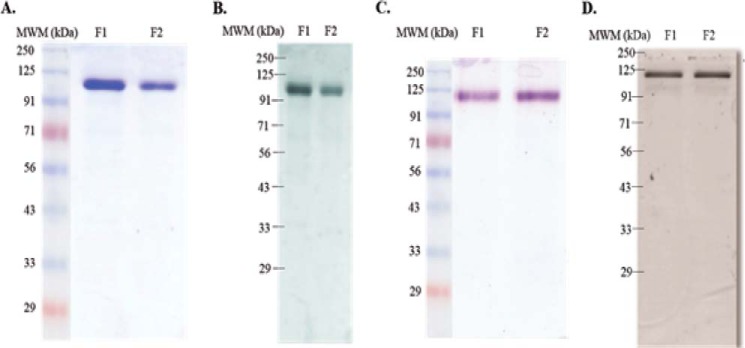
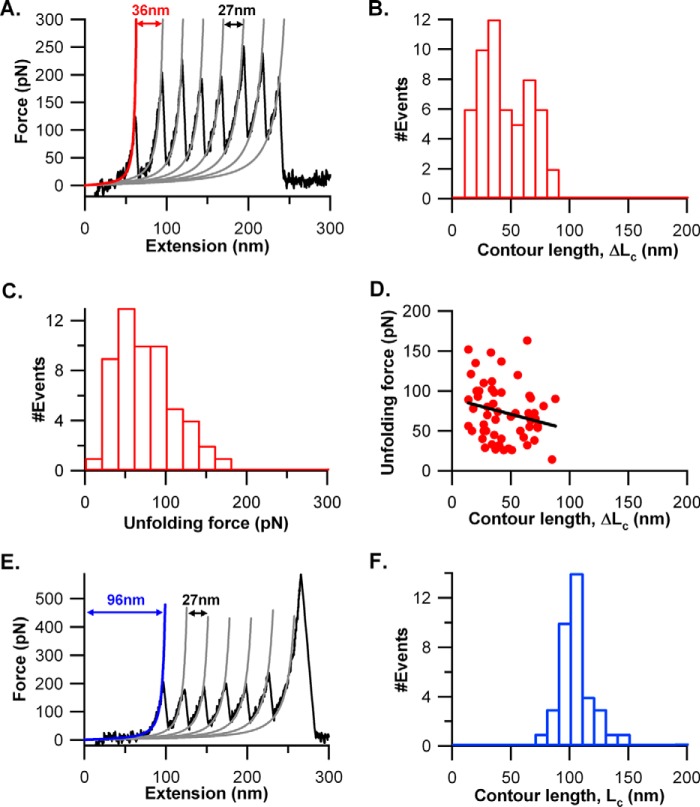
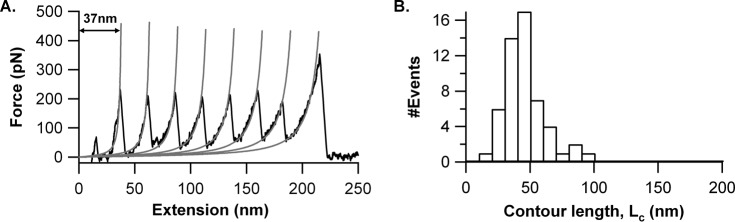
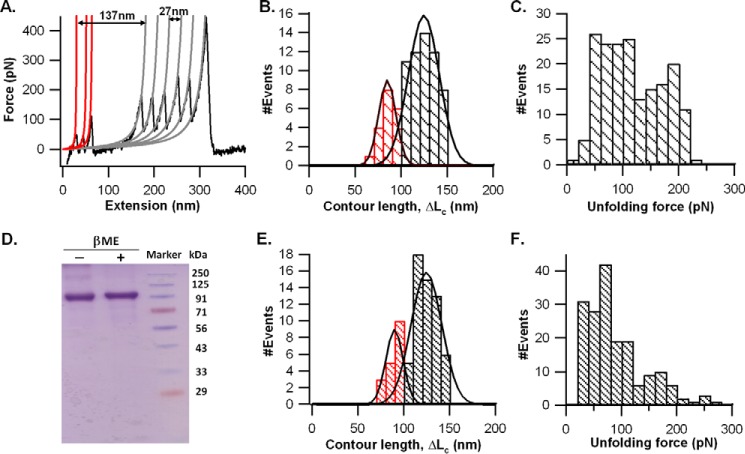
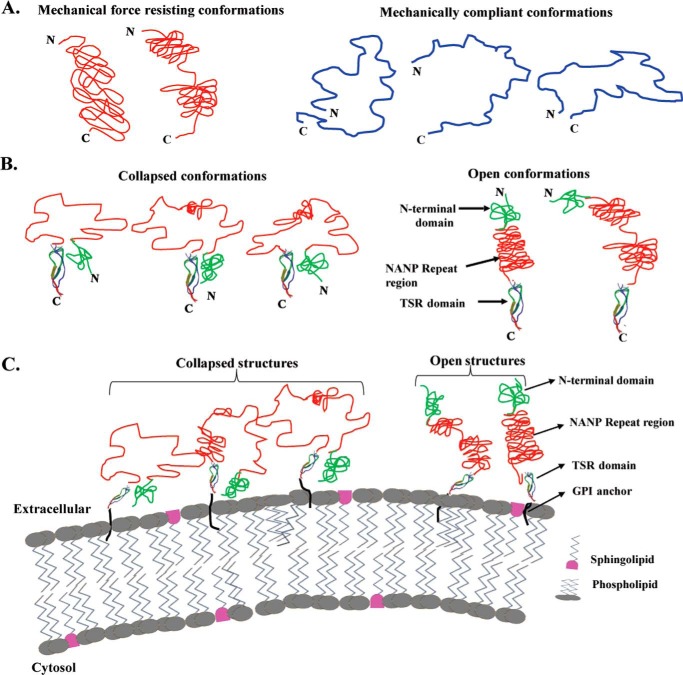
The most effective vaccine candidate of malaria is based on the Plasmodium falciparum circumsporozoite protein (CSP), a major surface protein implicated in the structural strength, motility, and immune evasion properties of the infective sporozoites. It is suspected that reversible conformational changes of CSP are required for infection of the mammalian host, but the detailed structure and dynamic properties of CSP remain incompletely understood, limiting our understanding of its function in the infection. Here, we report the structural and mechanical properties of the CSP studied using single-molecule force spectroscopy on several constructs, one including the central region of CSP, which is rich in NANP amino acid repeats (CSPrep), and a second consisting of a near full-length sequence without the signal and anchor hydrophobic domains (CSPΔHP). Our results show that the CSPrep is heterogeneous, with 40% of molecules requiring virtually no mechanical force to unfold (<10 piconewtons (pN)), suggesting that these molecules are mechanically compliant and perhaps act as entropic springs, whereas the remaining 60% are partially structured with low mechanical resistance (∼70 pN). CSPΔHP having multiple force peaks suggests specifically folded domains, with two major populations possibly indicating the open and collapsed forms. Our findings suggest that the overall low mechanical resistance of the repeat region, exposed on the outer surface of the sporozoites, combined with the flexible full-length conformations of CSP, may provide the sporozoites not only with immune evasion properties, but also with lubricating capacity required during its navigation through the mosquito and vertebrate host tissues. We anticipate that these findings would further assist in the design and development of future malarial vaccines.
Keywords: NANP; NANP repeats; Plasmodium falciparum; atomic force microscopy (AFM); circumsporozoite protein (CSP); force spectroscopy; malaria; malaria parasite; plasmodium; proline-rich peptides; proline-rich protein; single-molecule biophysics; sporozoite; structural biology.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- World Health Organization (2015) World Malaria Report 2015, WHO Publications, Geneva, Switzerland
-
- Cohen J., Nussenzweig V., Nussenzweig R., Vekemans J., and Leach A. (2010) From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum. Vaccin. 6, 90–96 - PubMed
-
- Kaslow D. C., and Biernaux S. (2015) RTS,S: toward a first landmark on the Malaria Vaccine Technology Roadmap. Vaccine 33, 7425–7432 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
