

Topoisomerase 3β is the major topoisomerase for mRNAs and linked to neurodevelopment and mental dysfunction
- PMID: 28039324
- PMCID: PMC5389537
- DOI: 10.1093/nar/gkw1293
Topoisomerase 3β is the major topoisomerase for mRNAs and linked to neurodevelopment and mental dysfunction
Abstract
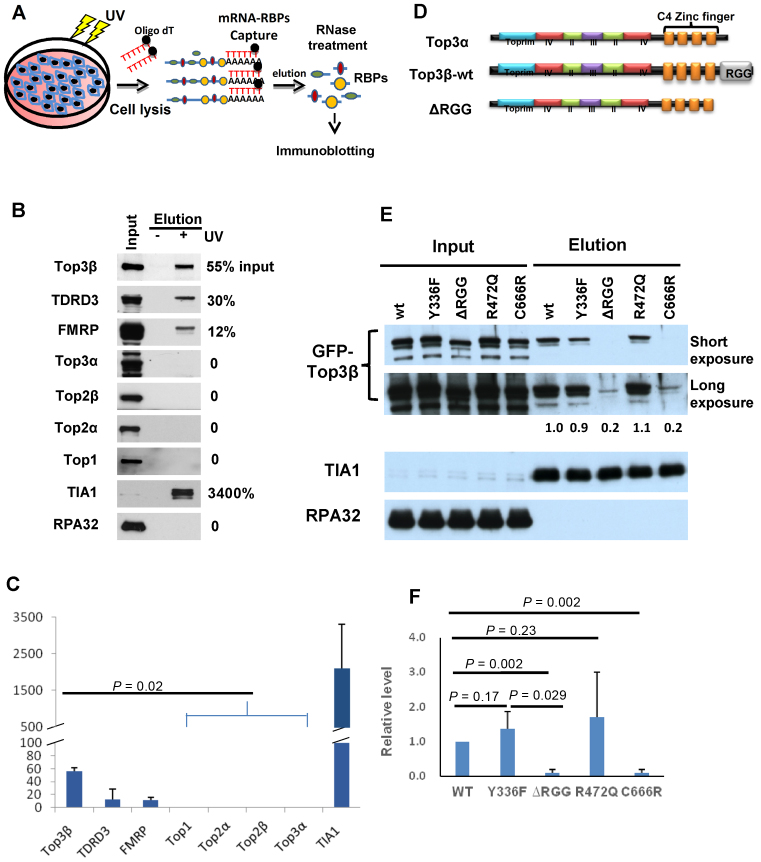
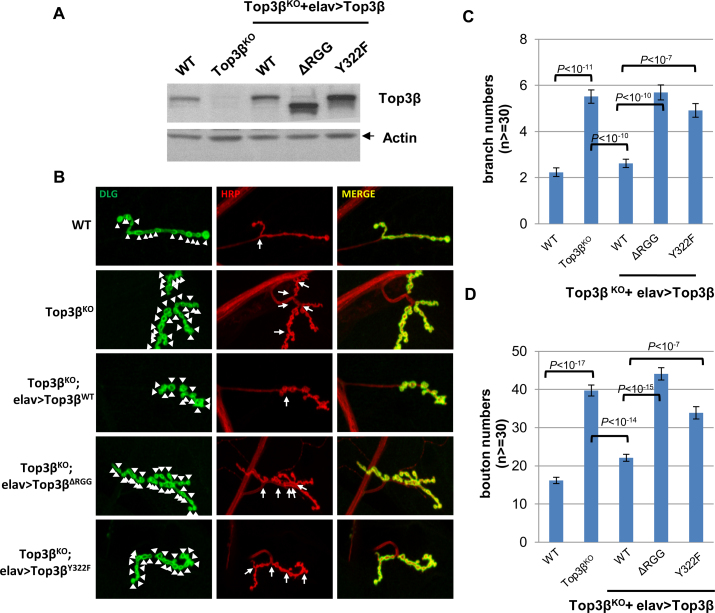
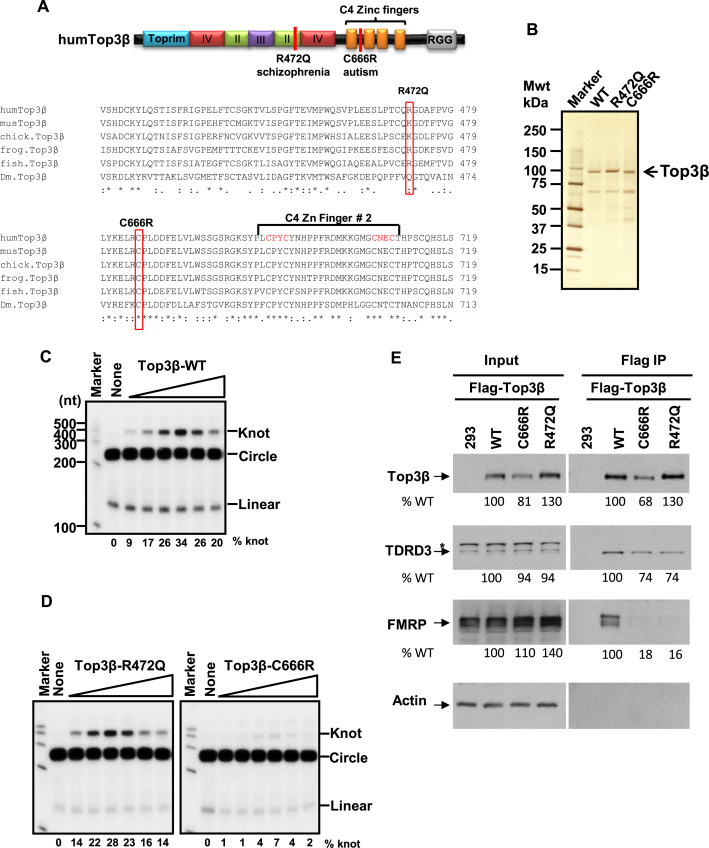
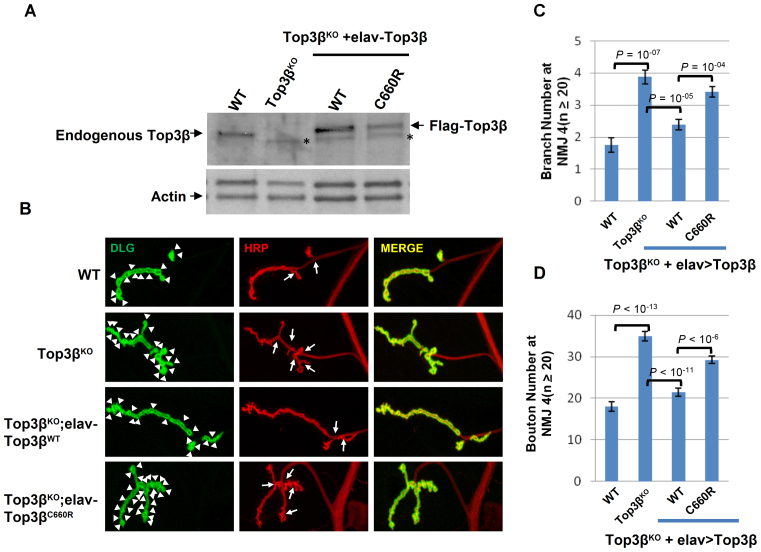
Human cells contain five topoisomerases in the nucleus and cytoplasm, but which one is the major topoisomerase for mRNAs is unclear. To date, Top3β is the only known topoisomerase that possesses RNA topoisomerase activity, binds mRNA translation machinery and interacts with an RNA-binding protein, FMRP, to promote synapse formation; and Top3β gene deletion has been linked to schizophrenia. Here, we show that Top3β is also the most abundant mRNA-binding topoisomerase in cells. Top3β, but not other topoisomerases, contains a distinctive RNA-binding domain; and deletion of this domain diminishes the amount of Top3β that associates with mRNAs, indicating that Top3β is specifically targeted to mRNAs by its RNA binding domain. Moreover, Top3β mutants lacking either its RNA-binding domain or catalytic residue fail to promote synapse formation, suggesting that Top3β requires both its mRNA-binding and catalytic activity to facilitate neurodevelopment. Notably, Top3β proteins bearing point mutations from schizophrenia and autism individuals are defective in association with FMRP; whereas one of the mutants is also deficient in binding mRNAs, catalyzing RNA topoisomerase reaction, and promoting synapse formation. Our data suggest that Top3β is the major topoisomerase for mRNAs, and requires both RNA binding and catalytic activity to promote neurodevelopment and prevent mental dysfunction.
Published by Oxford University Press on behalf of Nucleic Acids Research 2016.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
