

Inhibition of CRISPR-Cas9 with Bacteriophage Proteins
- PMID: 28041849
- PMCID: PMC5235966
- DOI: 10.1016/j.cell.2016.12.009
Inhibition of CRISPR-Cas9 with Bacteriophage Proteins
Abstract
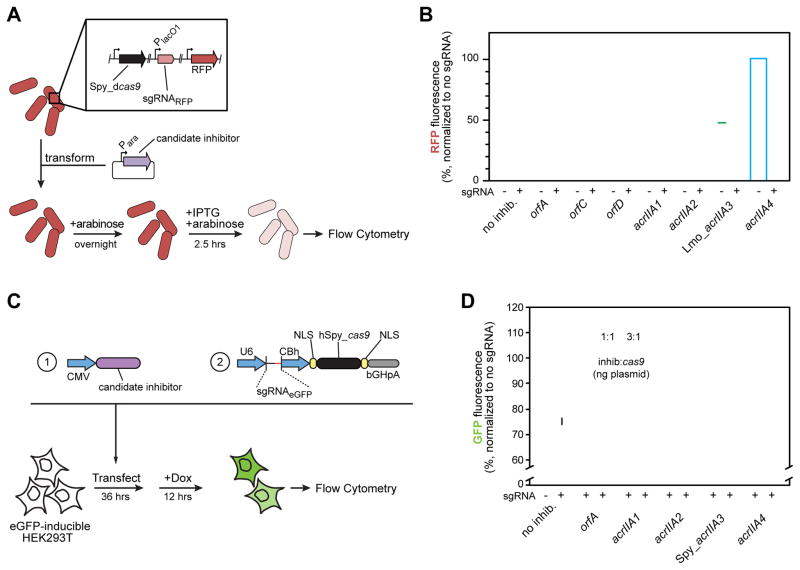
Bacterial CRISPR-Cas systems utilize sequence-specific RNA-guided nucleases to defend against bacteriophage infection. As a countermeasure, numerous phages are known that produce proteins to block the function of class 1 CRISPR-Cas systems. However, currently no proteins are known to inhibit the widely used class 2 CRISPR-Cas9 system. To find these inhibitors, we searched cas9-containing bacterial genomes for the co-existence of a CRISPR spacer and its target, a potential indicator for CRISPR inhibition. This analysis led to the discovery of four unique type II-A CRISPR-Cas9 inhibitor proteins encoded by Listeria monocytogenes prophages. More than half of L. monocytogenes strains with cas9 contain at least one prophage-encoded inhibitor, suggesting widespread CRISPR-Cas9 inactivation. Two of these inhibitors also blocked the widely used Streptococcus pyogenes Cas9 when assayed in Escherichia coli and human cells. These natural Cas9-specific "anti-CRISPRs" present tools that can be used to regulate the genome engineering activities of CRISPR-Cas9.
Keywords: CRISPR-Cas; Cas9; Cas9 inhibitor; Listeria monocytogenes; anti-CRISPR; bacteriophage; dCas9; gene editing; prophage.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
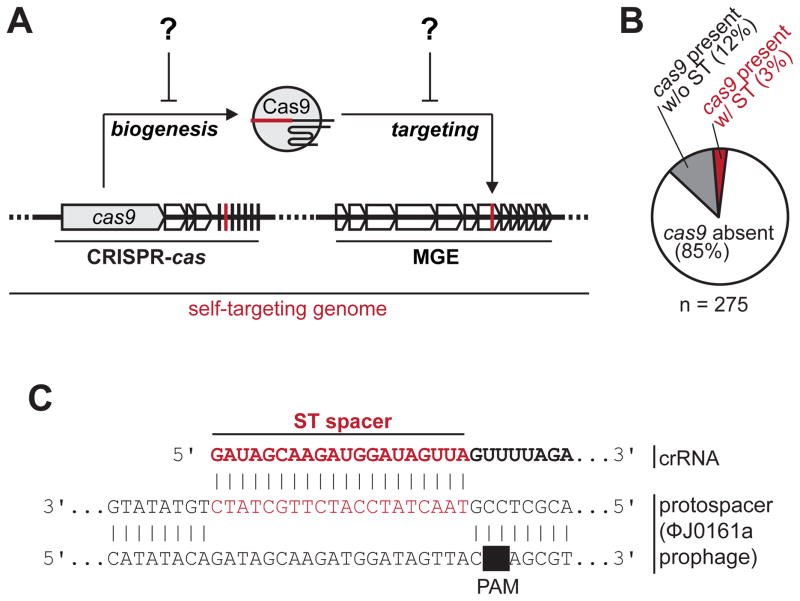
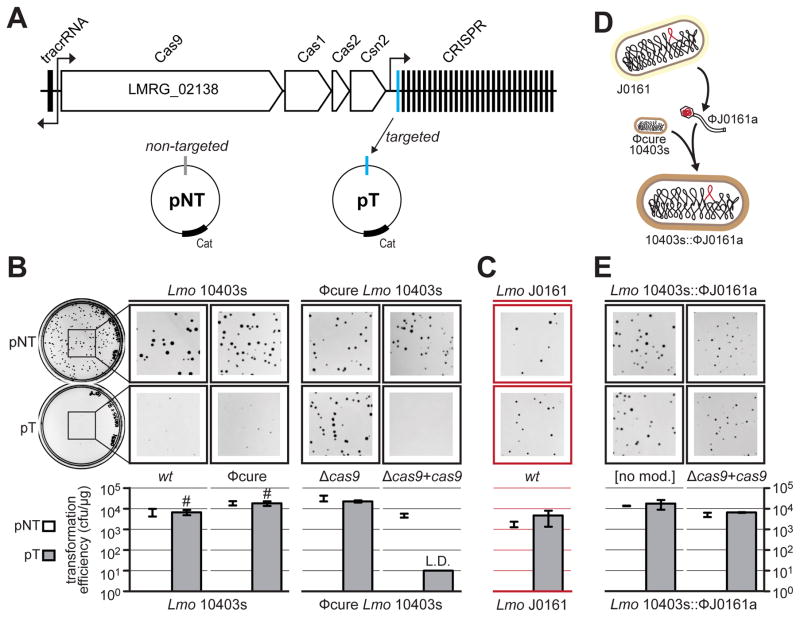
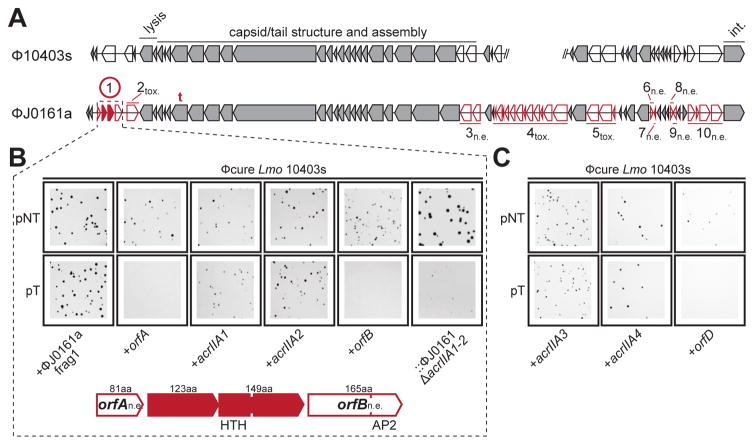

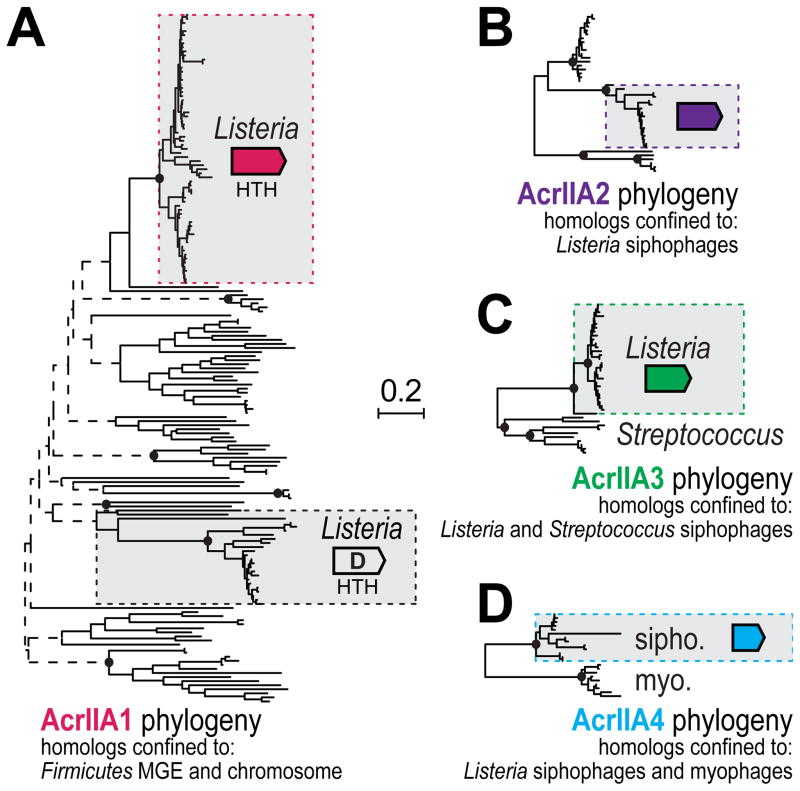
Comment in
-
The Interfaces of Genetic Conflict Are Hot Spots for Innovation.Cell. 2017 Jan 12;168(1-2):9-11. doi: 10.1016/j.cell.2016.12.007. Epub 2017 Jan 12. Cell. 2017. PMID: 28086100 Free PMC article.
References
-
- Barrangou R, Doudna JA. Applications of CRISPR technologies in research and beyond. Nature Biotechnology. 2016;34:933–941. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
