

SREBP1 Contributes to Resolution of Pro-inflammatory TLR4 Signaling by Reprogramming Fatty Acid Metabolism
- PMID: 28041958
- PMCID: PMC5568699
- DOI: 10.1016/j.cmet.2016.11.009
SREBP1 Contributes to Resolution of Pro-inflammatory TLR4 Signaling by Reprogramming Fatty Acid Metabolism
Abstract
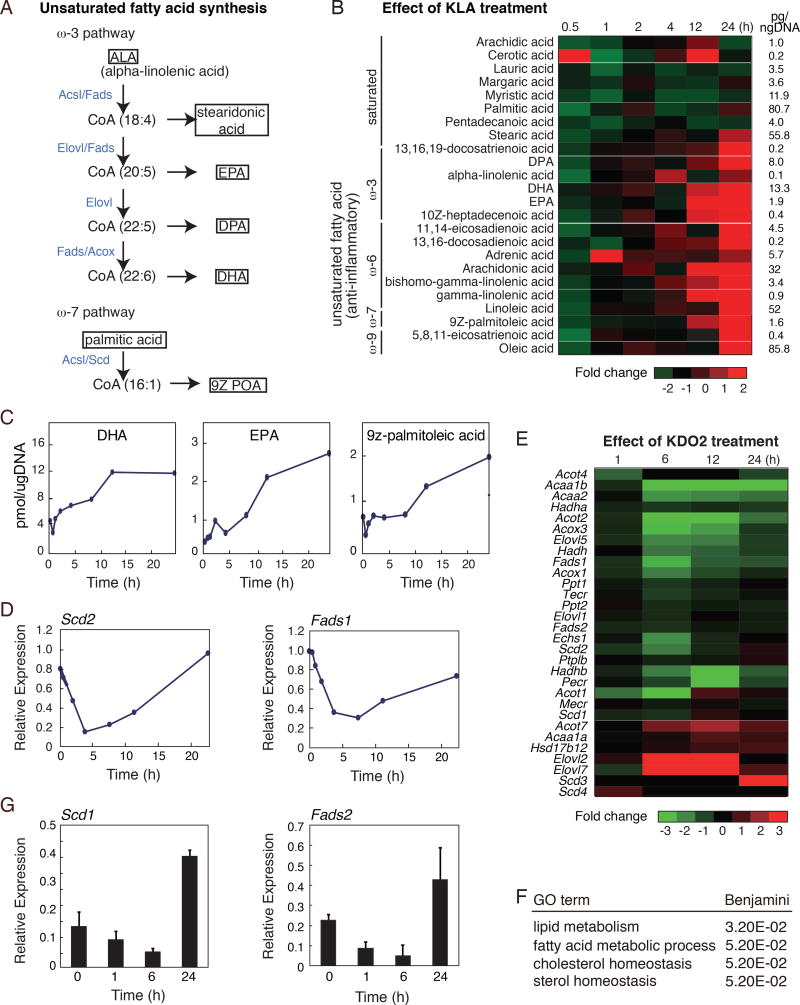
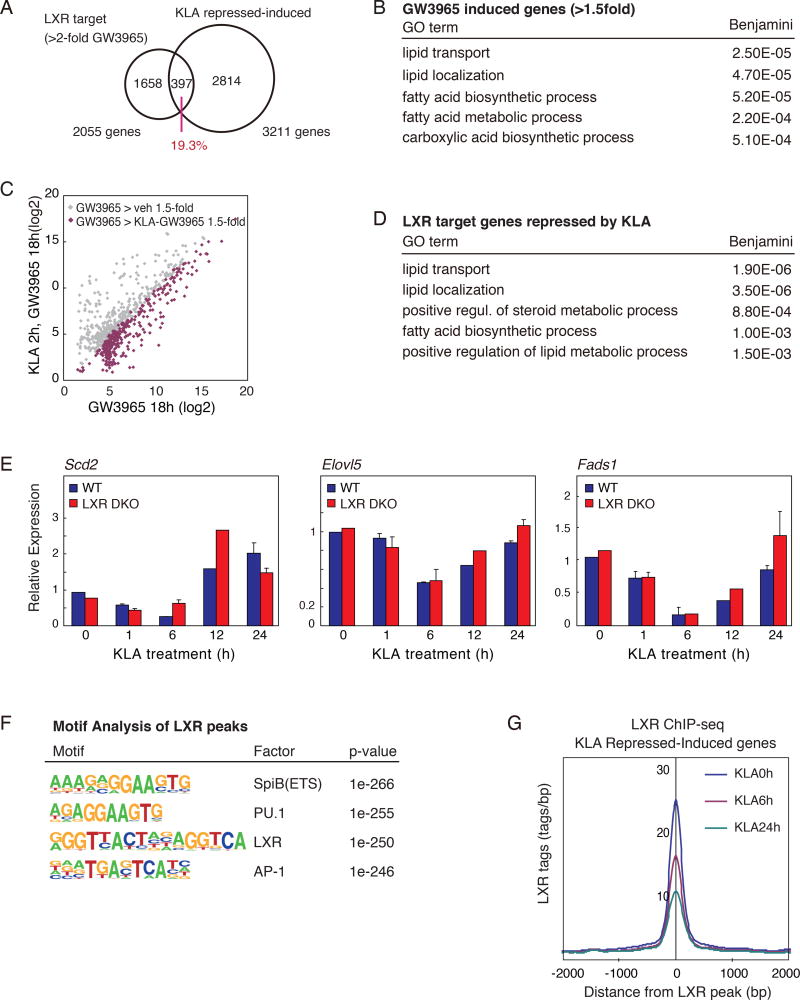
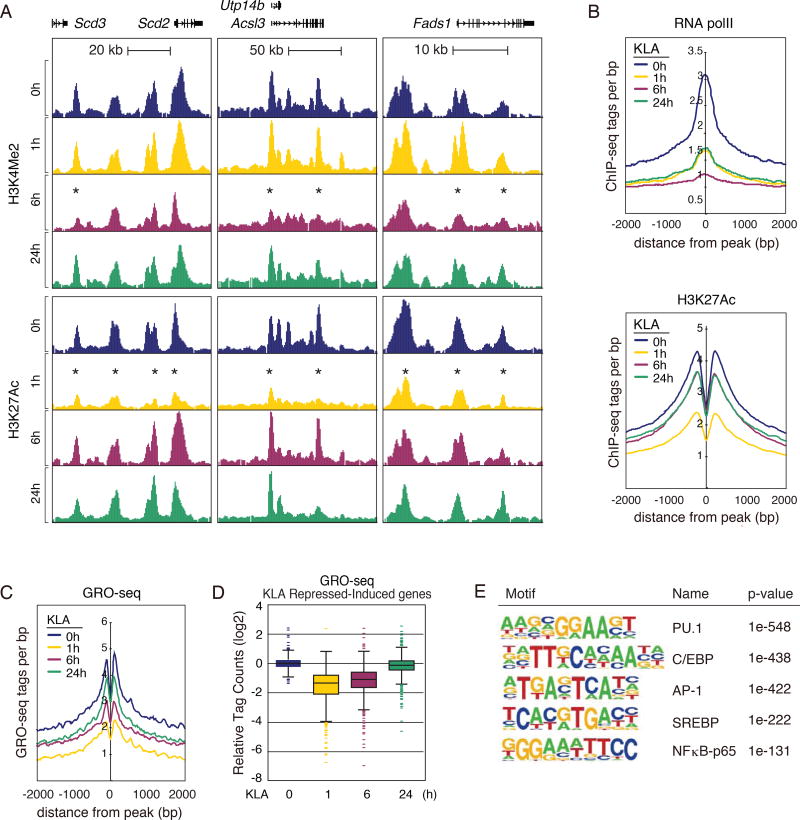
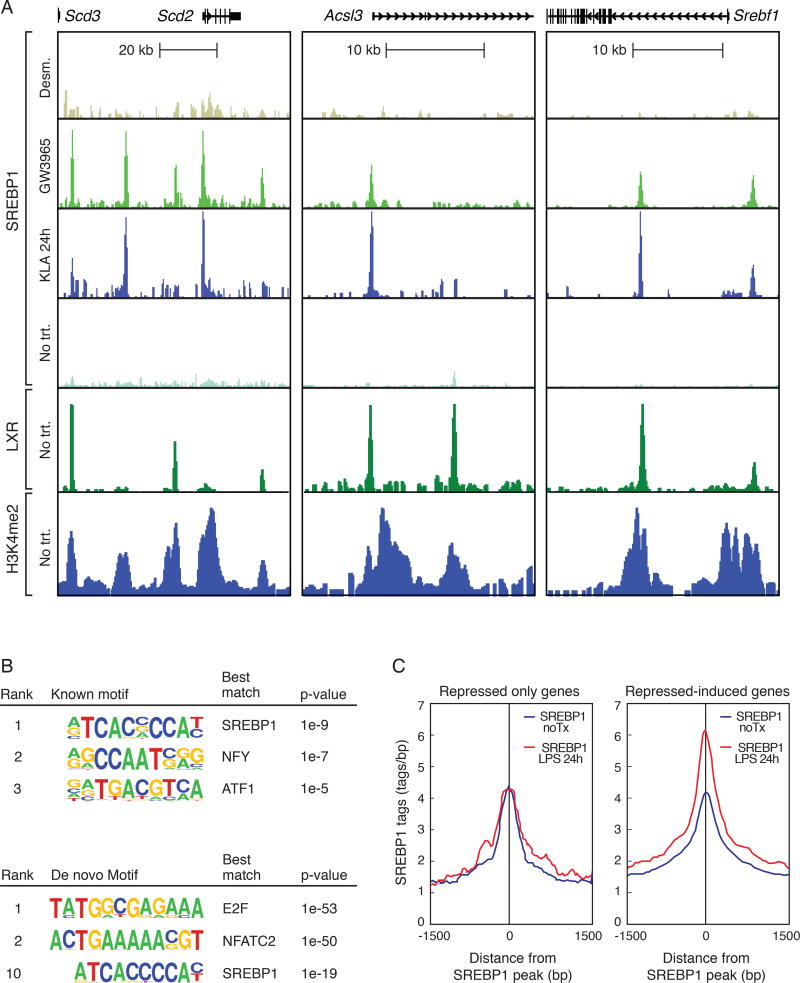
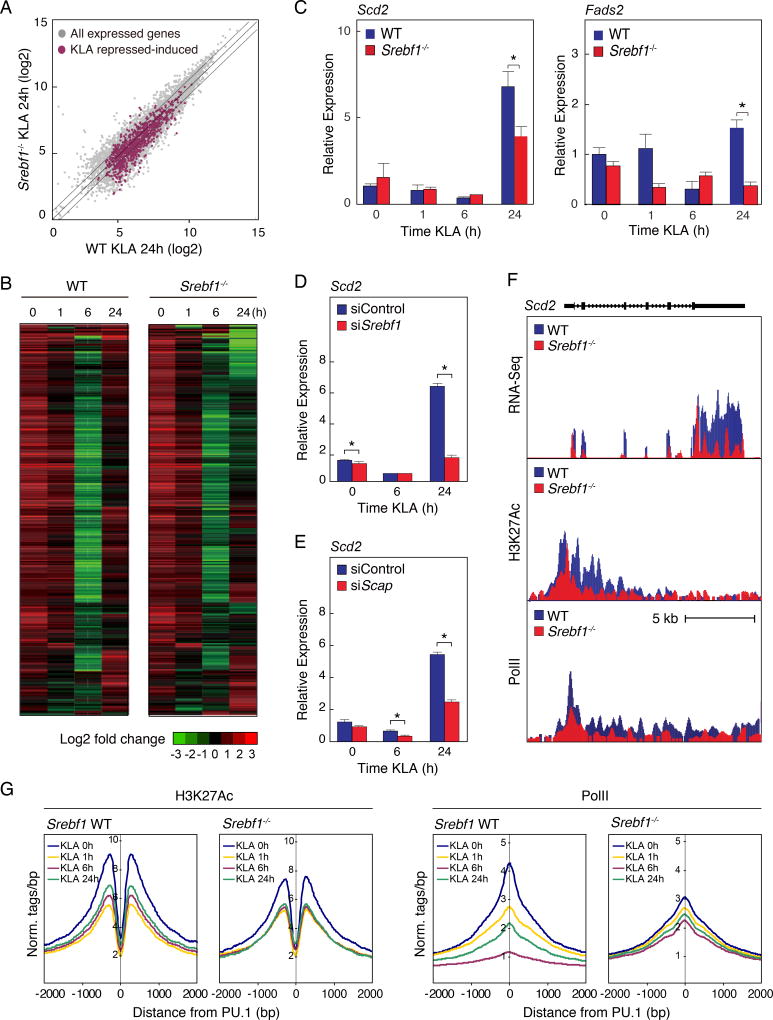
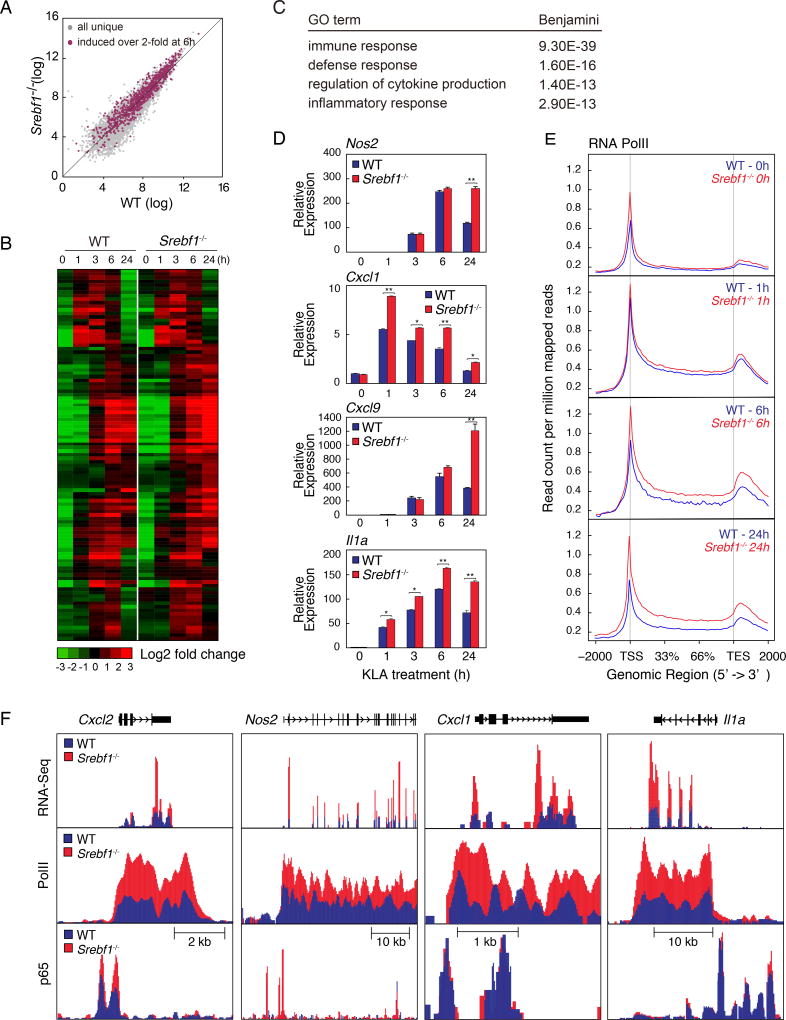
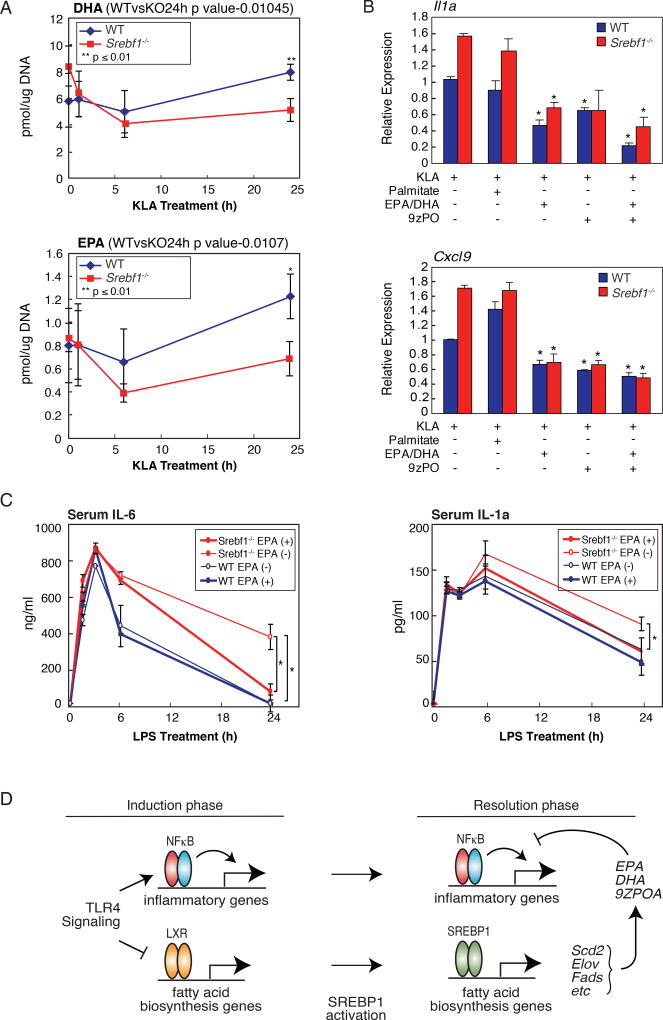
Macrophages play pivotal roles in both the induction and resolution phases of inflammatory processes. Macrophages have been shown to synthesize anti-inflammatory fatty acids in an LXR-dependent manner, but whether the production of these species contributes to the resolution phase of inflammatory responses has not been established. Here, we identify a biphasic program of gene expression that drives production of anti-inflammatory fatty acids 12-24 hr following TLR4 activation and contributes to downregulation of mRNAs encoding pro-inflammatory mediators. Unexpectedly, rather than requiring LXRs, this late program of anti-inflammatory fatty acid biosynthesis is dependent on SREBP1 and results in the uncoupling of NFκB binding from gene activation. In contrast to previously identified roles of SREBP1 in promoting production of IL1β during the induction phase of inflammation, these studies provide evidence that SREBP1 also contributes to the resolution phase of TLR4-induced gene activation by reprogramming macrophage lipid metabolism.
Keywords: DHA; EPA; SREBP1; fatty acid metabolism; inflammation; innate immunity; lipid metabolism; resolution; transcriptional regulation; unsaturated fatty acids.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Castrillo A, Joseph SB, Vaidya SA, Haberland M, Fogelman AM, Cheng G, Tontonoz P. Crosstalk between LXR and toll-like receptor signaling mediates bacterial and viral antagonism of cholesterol metabolism. Mol Cell. 2003;12:805–816. - PubMed
-
- Cildir G, Akincilar SC, Tergaonkar V. Chronic adipose tissue inflammation: all immune cells on the stage. Trends Mol Med. 2013;19:487–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
