

Aquaporin-Mediated Water and Hydrogen Peroxide Transport Is Involved in Normal Human Spermatozoa Functioning
- PMID: 28042826
- PMCID: PMC5297701
- DOI: 10.3390/ijms18010066
Aquaporin-Mediated Water and Hydrogen Peroxide Transport Is Involved in Normal Human Spermatozoa Functioning
Abstract
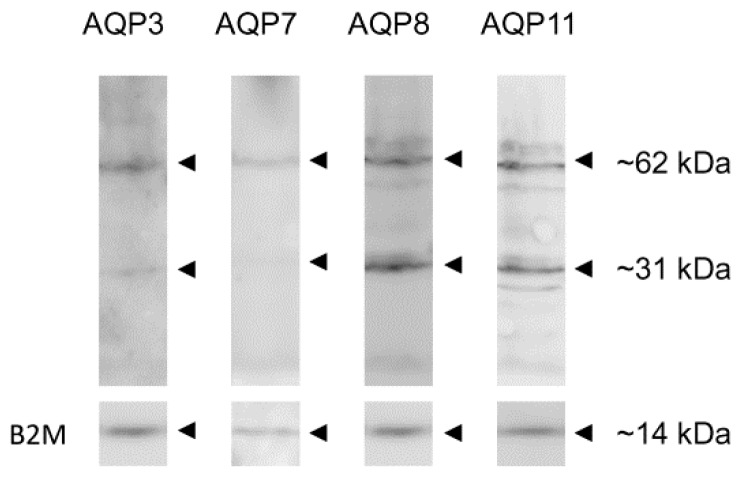
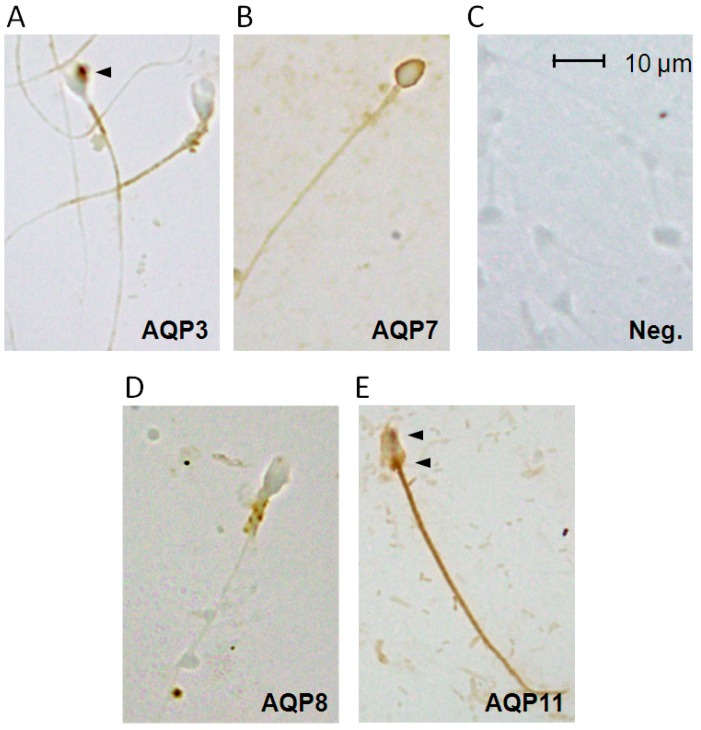
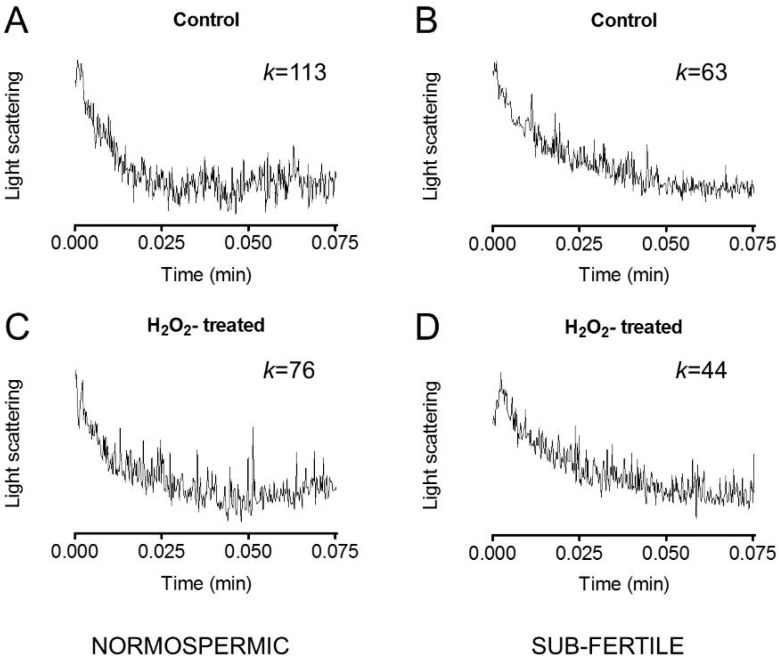
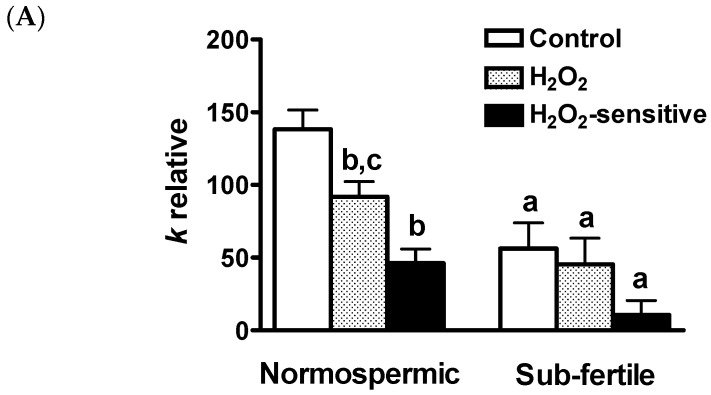
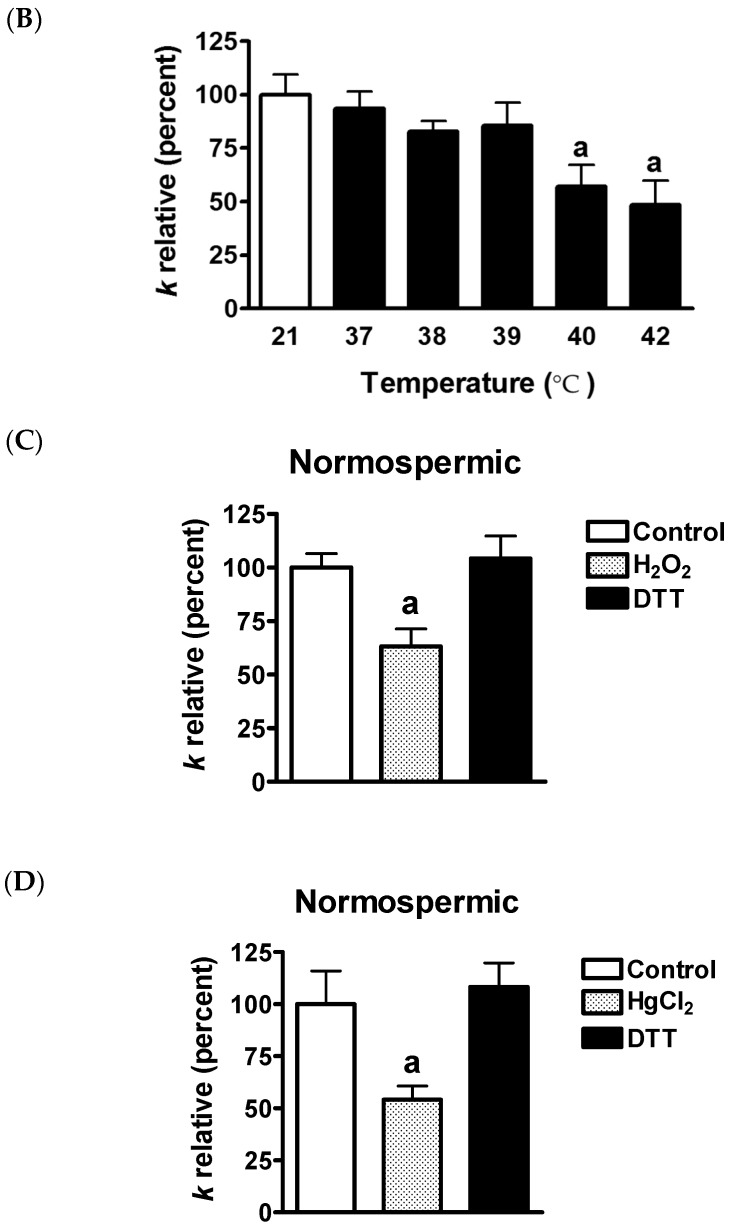
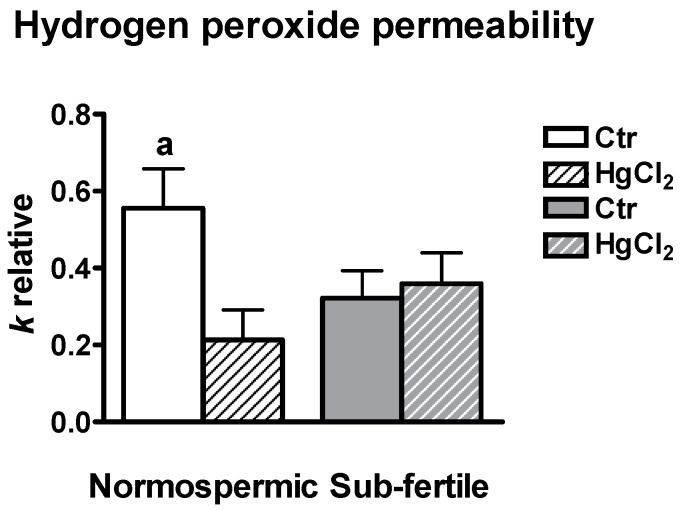
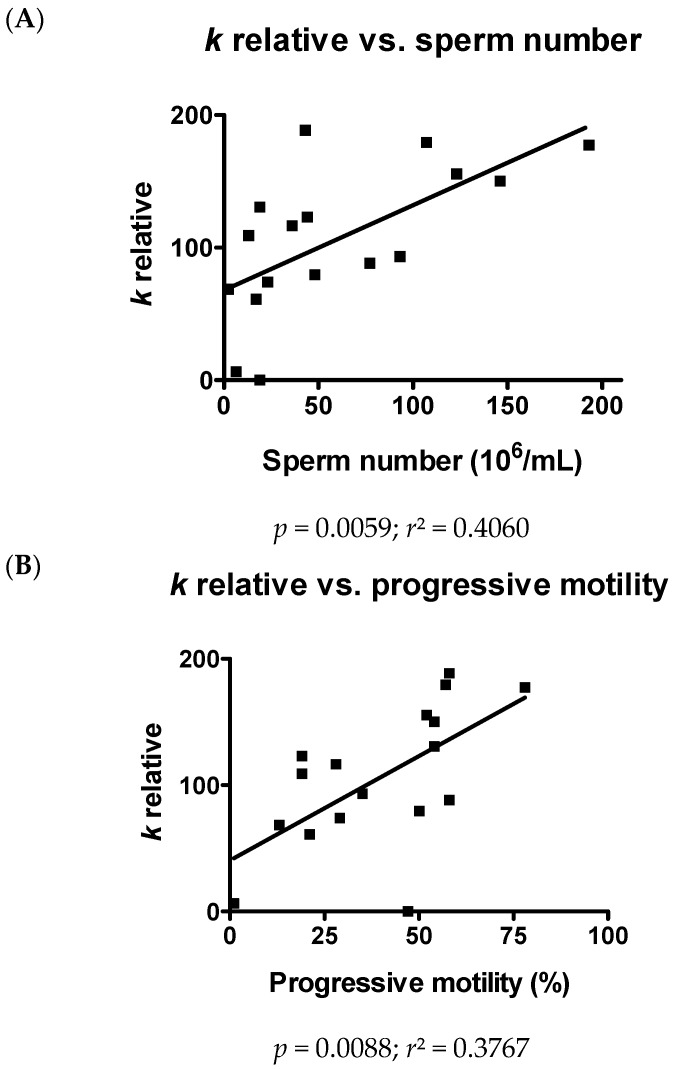
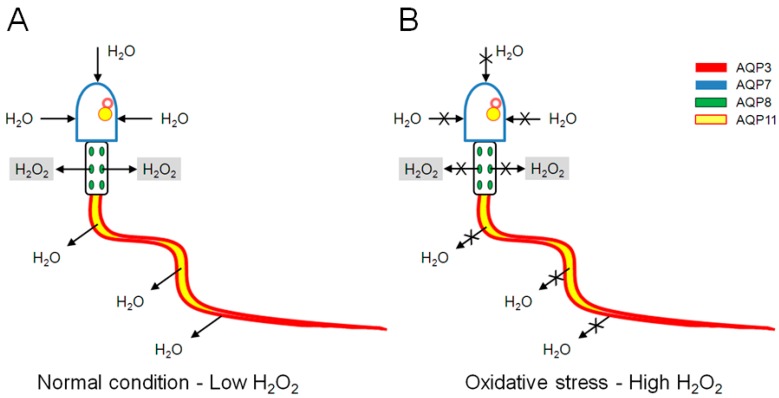
Different aquaporins (AQPs) are expressed in human sperm cells and with a different localization. Their function has been related to cell volume control in response to the osmotic changes encountered passing from the epididymal fluid to the cervical mucus or involved in the end stage of cytoplasm removal during sperm maturation. Recently, AQPs have also shown hydrogen peroxide (H₂O₂) permeability properties. Here, we investigate the expression, localization and functioning of AQPs in human sperm cells with particular attention to their role as peroxiporins in reactive oxygen species (ROS) scavenging in both normospermic and sub-fertile human subjects. Western blotting and immunocytochemistry were used to confirm and clarify the AQPs expression and localization. Water and H₂O₂ permeability was tested by stopped flow light scattering method and by the CM-H2DCFDA (5-(and-6)-chloromethyl-2',7'-dichlorodihydro-fluorescein diacetate, acetyl ester) H₂O₂ fluorescence probe, respectively. AQP3, -7, -8, and -11 proteins were found in human sperm cells and localized in the head (AQP7), in the middle piece (AQP8) and in the tail (AQP3 and -11) in both the plasma membrane and in intracellular structures. Sperm cells showed water and H₂O₂ permeability which was reversibly inhibited by H₂O₂, heat stress and the AQP inhibitor HgCl₂. Reduced functionality was observed in patients with compromised basal semen parameters. Present findings suggest that AQPs are involved in both volume regulation and ROS elimination. The relationship between sperm number and motility and AQP functioning was also demonstrated.
Keywords: aquaporins-7; aquaporins-8; oxidative stress; sperm motility; sterility; water channel.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- De Lamirande E., Gagnon C. Reactive oxygen species and human spermatozoa. I. Effects on the motility of intact spermatozoa and on sperm axonemes. J. Androl. 1992;13:368–378. - PubMed
-
- De Lamirande E., Gagnon C. Reactive oxygen species and human spermatozoa. II. Depletion of adenosine triphosphate plays an important role in the inhibition of sperm motility. J. Androl. 1992;13:379–386. - PubMed
-
- De Lamirande E., Eiley D., Gagnon C. Inverse relationship between the induction of human sperm capacitation and spontaneous acrosome reaction by various biological fluids and the superoxide scavenging capacity of these fluids. Int. J. Androl. 1993;16:258–266. doi: 10.1111/j.1365-2605.1993.tb01189.x. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
