

Spatial and temporal signal processing and decision making by MAPK pathways
- PMID: 28043970
- PMCID: PMC5294789
- DOI: 10.1083/jcb.201609124
Spatial and temporal signal processing and decision making by MAPK pathways
Abstract
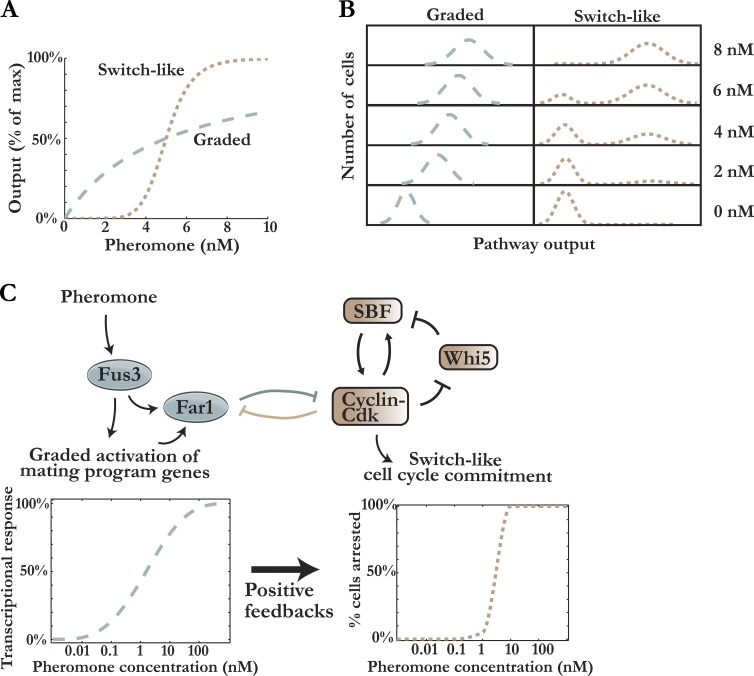
Mitogen-activated protein kinase (MAPK) pathways are conserved from yeast to man and regulate a variety of cellular processes, including proliferation and differentiation. Recent developments show how MAPK pathways perform exquisite spatial and temporal signal processing and underscores the importance of studying the dynamics of signaling pathways to understand their physiological response. The importance of dynamic mechanisms that process input signals into graded downstream responses has been demonstrated in the pheromone-induced and osmotic stress-induced MAPK pathways in yeast and in the mammalian extracellular signal-regulated kinase MAPK pathway. Particularly, recent studies in the yeast pheromone response have shown how positive feedback generates switches, negative feedback enables gradient detection, and coherent feedforward regulation underlies cellular memory. More generally, a new wave of quantitative single-cell studies has begun to elucidate how signaling dynamics determine cell physiology and represents a paradigm shift from descriptive to predictive biology.
© 2017 Atay and Skotheim.
Figures




References
-
- Amir S.M., Carraway T.F. Jr., Kohn L.D., and Winand R.J.. 1973. The binding of thyrotropin to isolated bovine thyroid plasma membranes. J. Biol. Chem. 248:4092–4100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
