

Mechanosensing by the nucleus: From pathways to scaling relationships
- PMID: 28043971
- PMCID: PMC5294790
- DOI: 10.1083/jcb.201610042
Mechanosensing by the nucleus: From pathways to scaling relationships
Abstract
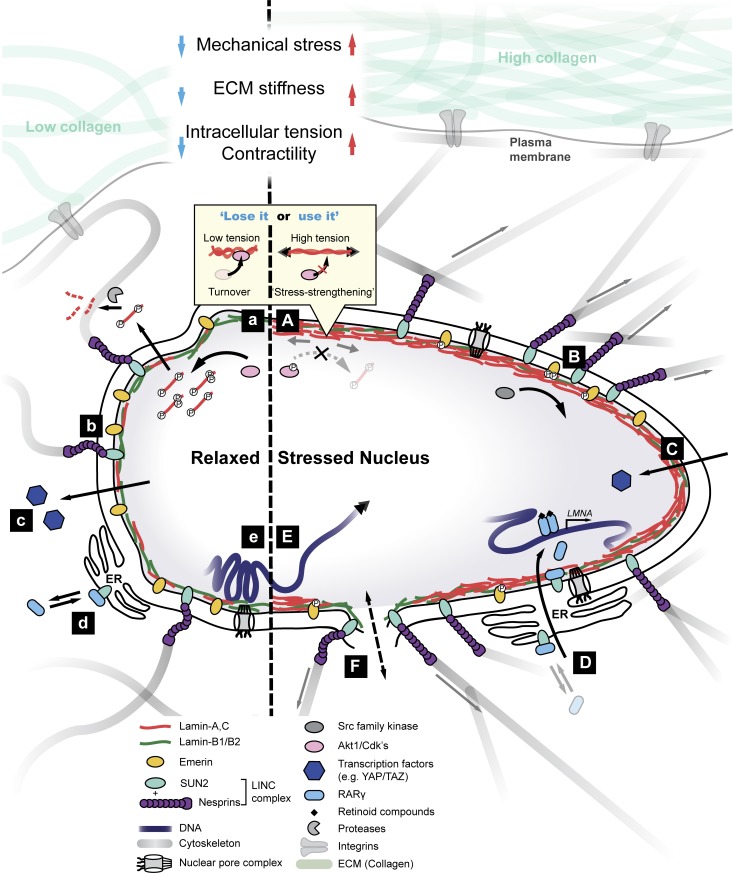
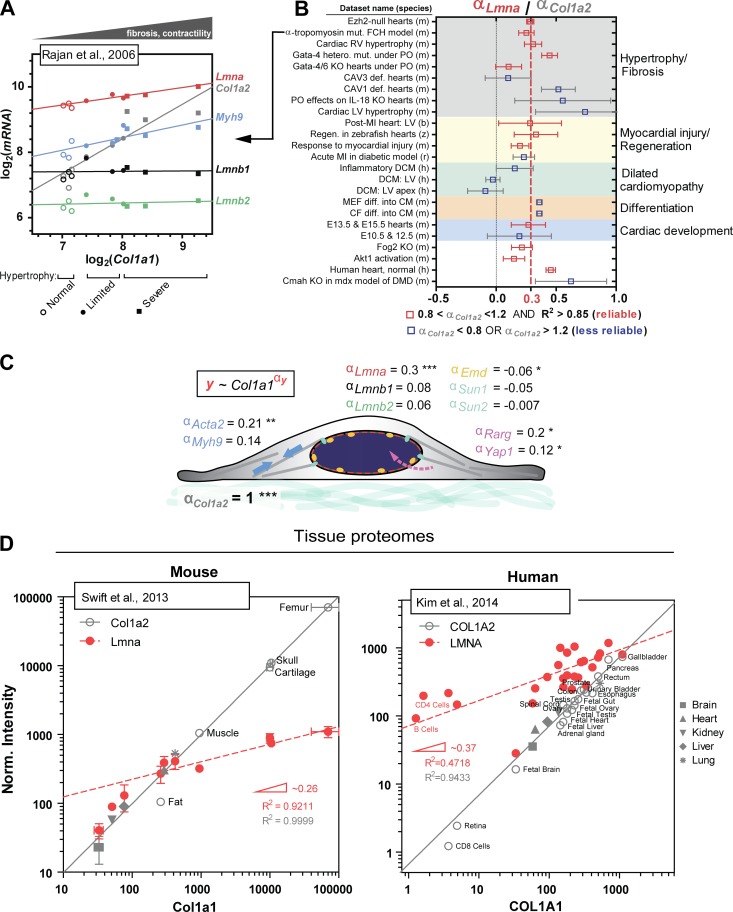
The nucleus is linked mechanically to the extracellular matrix via multiple polymers that transmit forces to the nuclear envelope and into the nuclear interior. Here, we review some of the emerging mechanisms of nuclear mechanosensing, which range from changes in protein conformation and transcription factor localization to chromosome reorganization and membrane dilation up to rupture. Nuclear mechanosensing encompasses biophysically complex pathways that often converge on the main structural proteins of the nucleus, the lamins. We also perform meta-analyses of public transcriptomics and proteomics data, which indicate that some of the mechanosensing pathways relaying signals from the collagen matrix to the nucleus apply to a broad range of species, tissues, and diseases.
© 2017 Cho et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
