

The Pseudomonas putida T6SS is a plant warden against phytopathogens
- PMID: 28045455
- PMCID: PMC5363822
- DOI: 10.1038/ismej.2016.169
The Pseudomonas putida T6SS is a plant warden against phytopathogens
Abstract
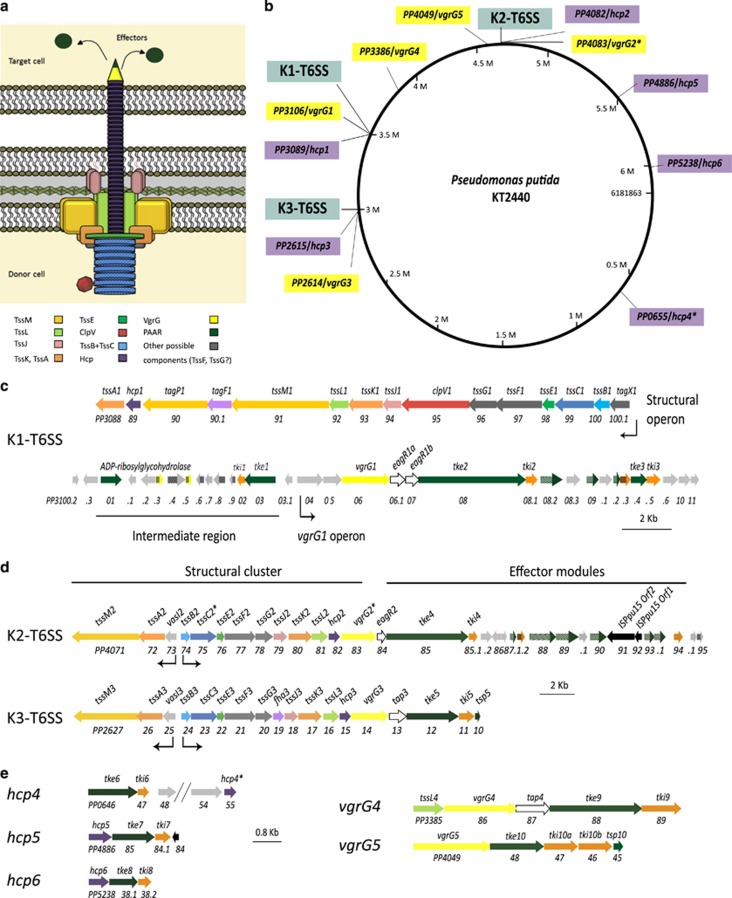
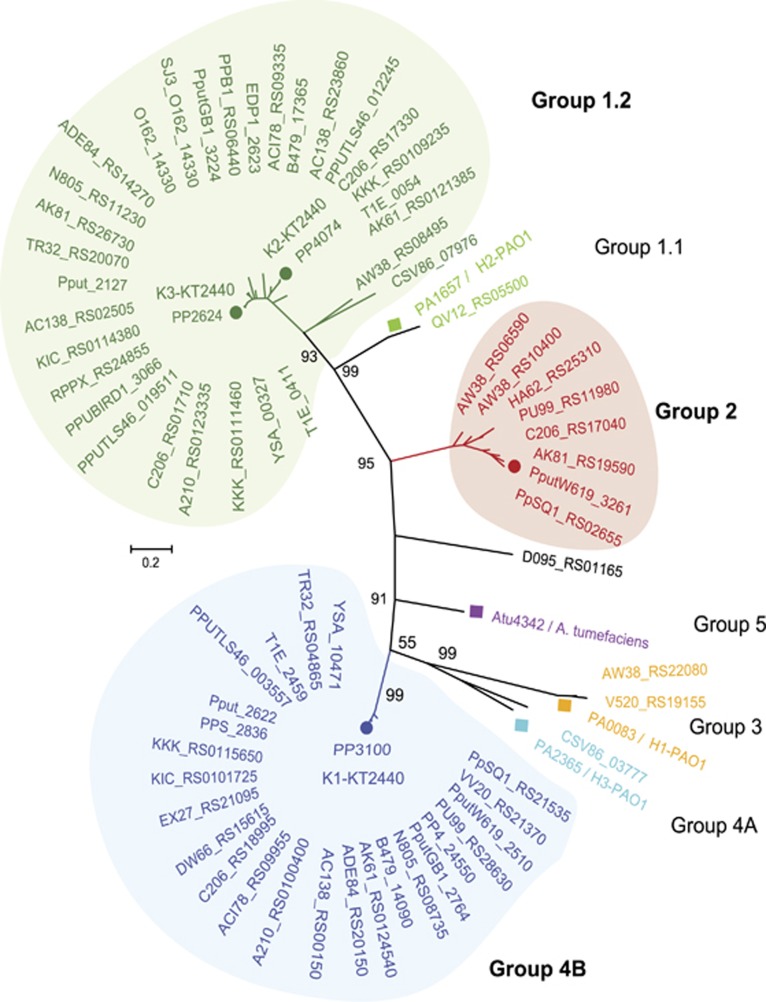
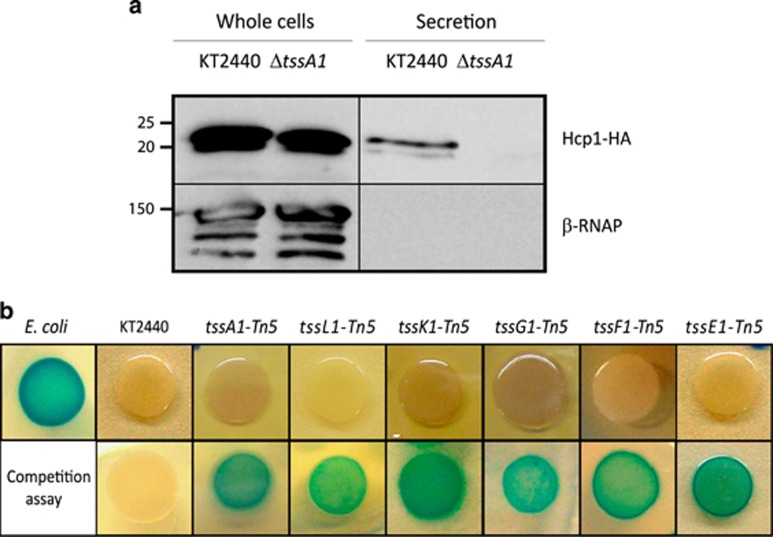
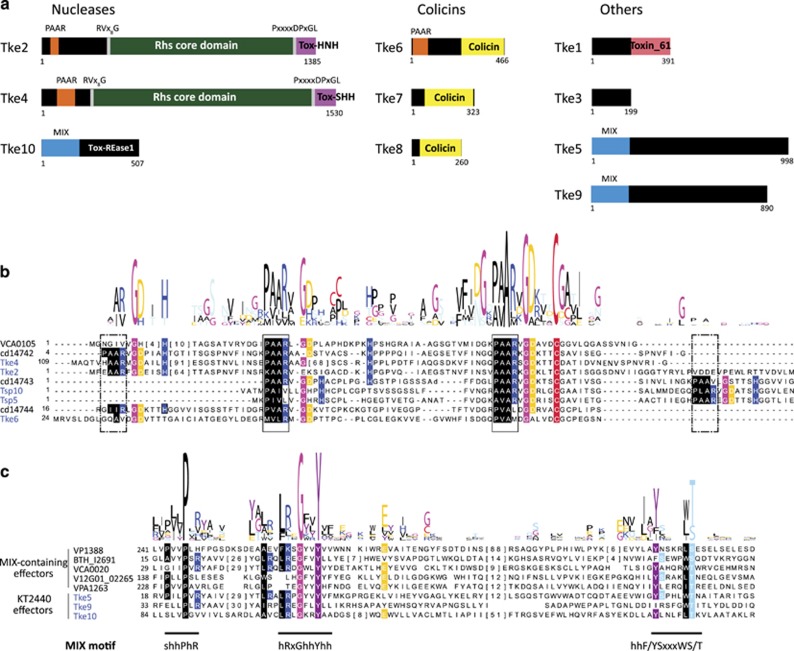
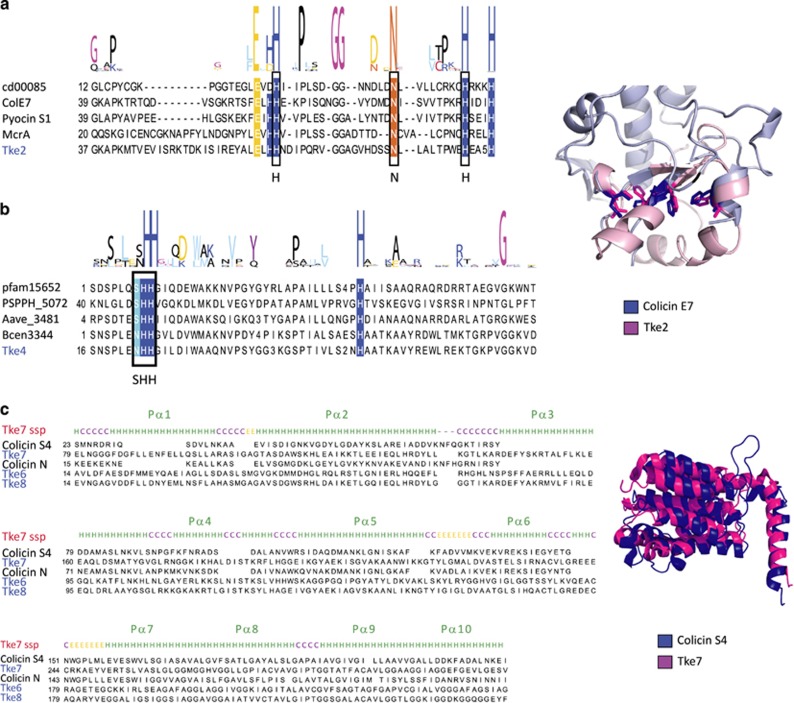
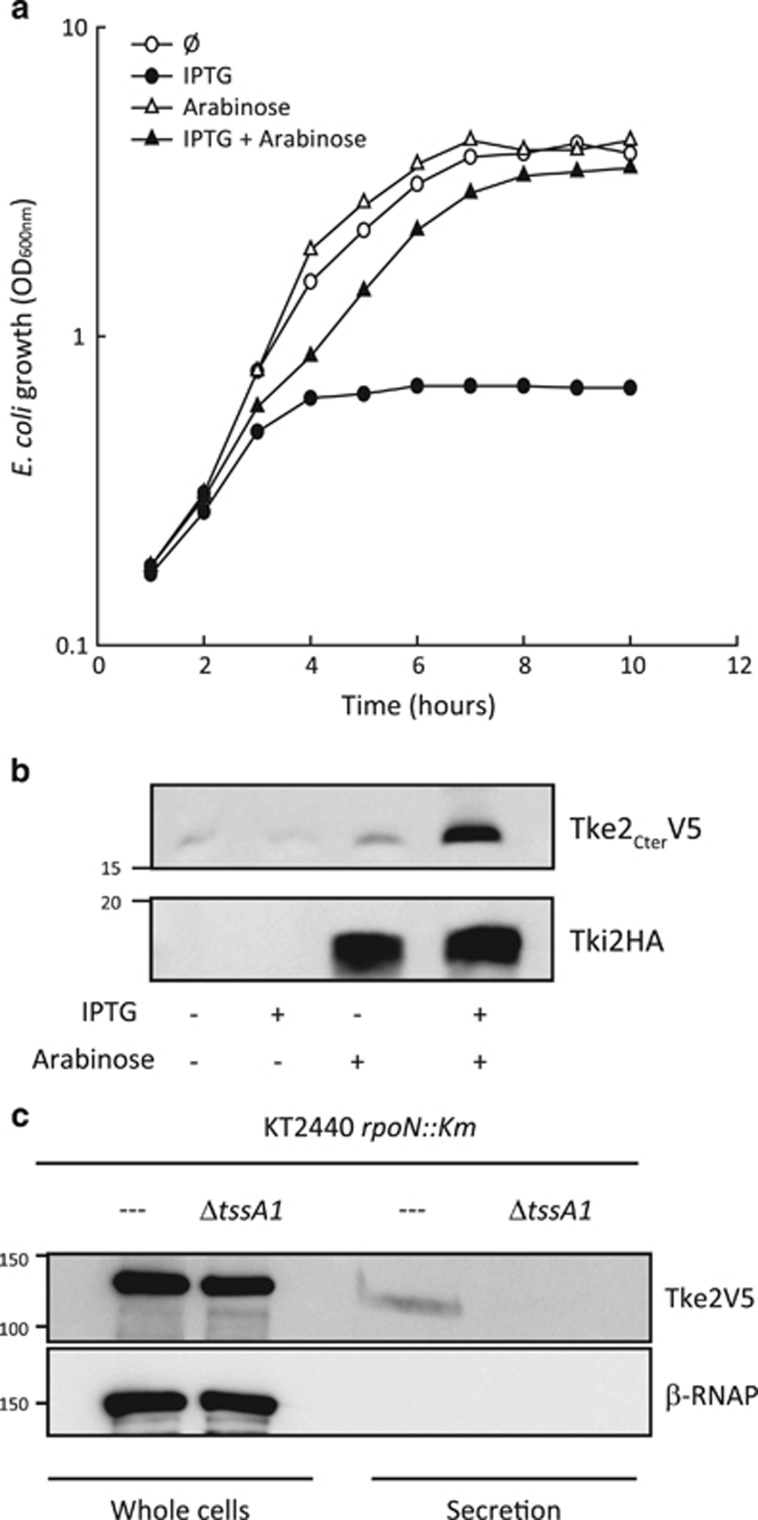
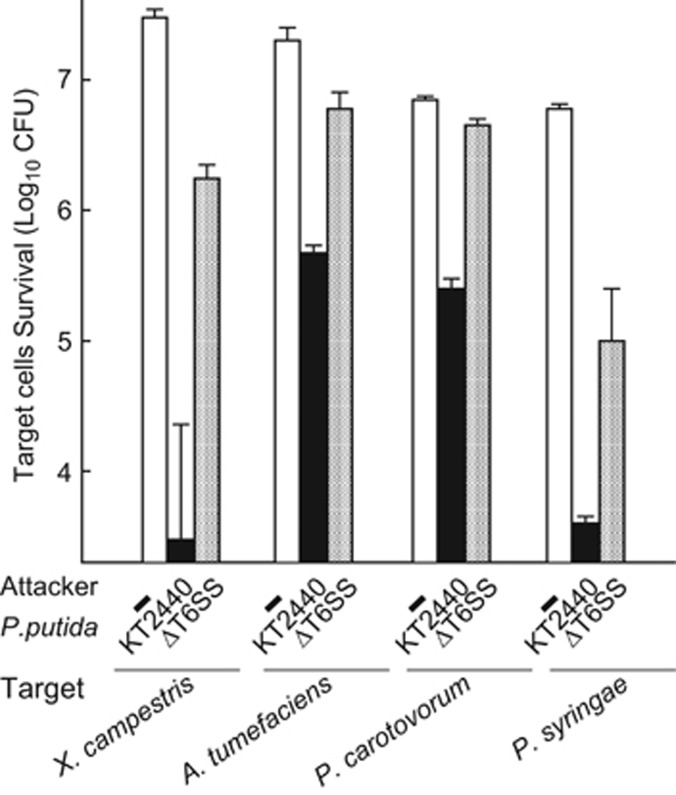
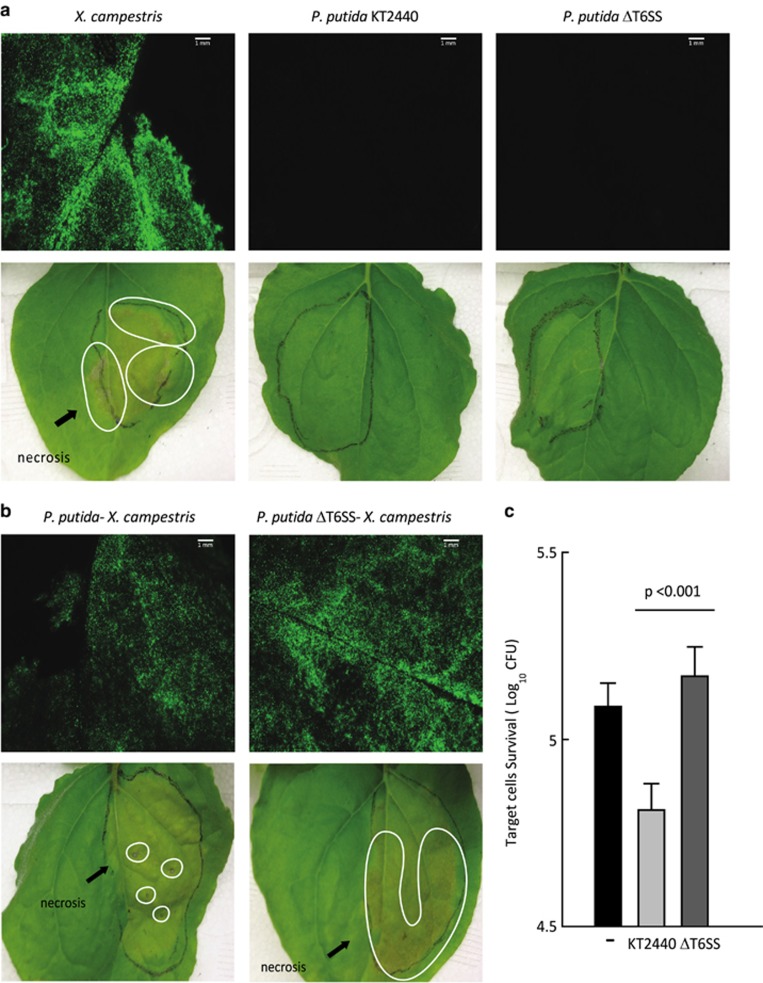
Bacterial type VI secretion systems (T6SSs) are molecular weapons designed to deliver toxic effectors into prey cells. These nanomachines have an important role in inter-bacterial competition and provide advantages to T6SS active strains in polymicrobial environments. Here we analyze the genome of the biocontrol agent Pseudomonas putida KT2440 and identify three T6SS gene clusters (K1-, K2- and K3-T6SS). Besides, 10 T6SS effector-immunity pairs were found, including putative nucleases and pore-forming colicins. We show that the K1-T6SS is a potent antibacterial device, which secretes a toxic Rhs-type effector Tke2. Remarkably, P. putida eradicates a broad range of bacteria in a K1-T6SS-dependent manner, including resilient phytopathogens, which demonstrates that the T6SS is instrumental to empower P. putida to fight against competitors. Furthermore, we observed a drastically reduced necrosis on the leaves of Nicotiana benthamiana during co-infection with P. putida and Xanthomonas campestris. Such protection is dependent on the activity of the P. putida T6SS. Many routes have been explored to develop biocontrol agents capable of manipulating the microbial composition of the rhizosphere and phyllosphere. Here we unveil a novel mechanism for plant biocontrol, which needs to be considered for the selection of plant wardens whose mission is to prevent phytopathogen infections.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Amer GA, Utkhede RS. (2000). Development of formulations of biological agents for management of root rot of lettuce and cucumber. Can J Microbiol 46: 809–816. - PubMed
-
- Aschtgen M-SS, Thomas MS, Cascales E. (2010). Anchoring the type VI secretion system to the peptidoglycan: TssL, TagL, TagP... what else? Virulence 1: 535–540. - PubMed
