

Homologous Recombination Defective Arabidopsis Mutants Exhibit Enhanced Sensitivity to Abscisic Acid
- PMID: 28046013
- PMCID: PMC5207409
- DOI: 10.1371/journal.pone.0169294
Homologous Recombination Defective Arabidopsis Mutants Exhibit Enhanced Sensitivity to Abscisic Acid
Abstract
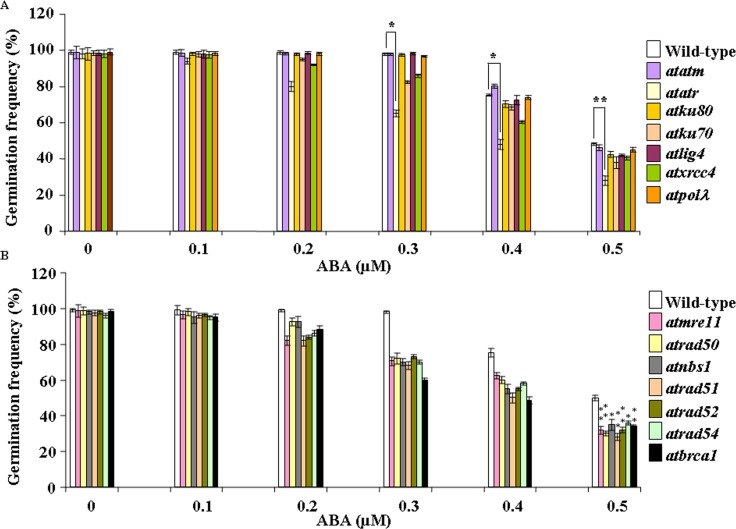
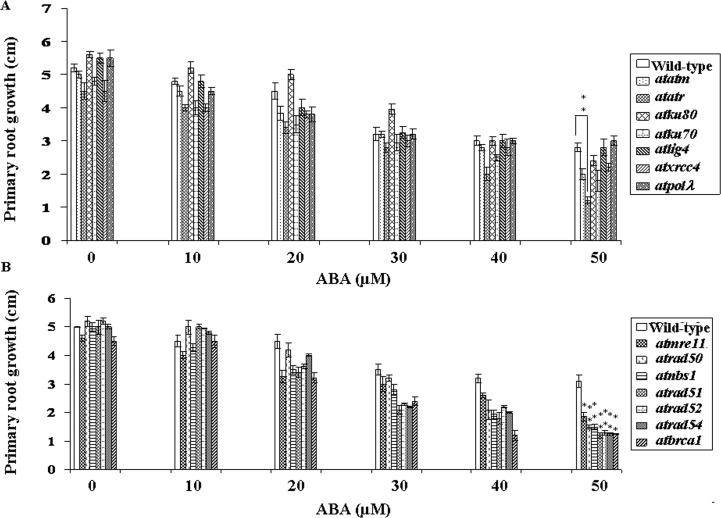
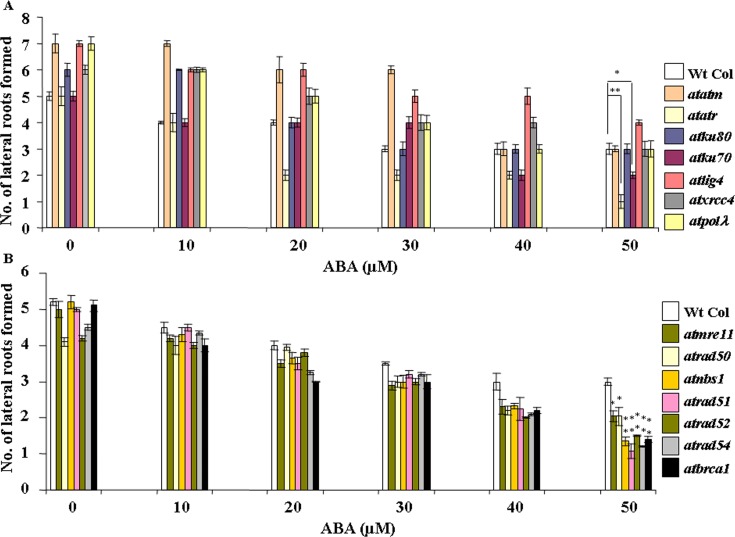
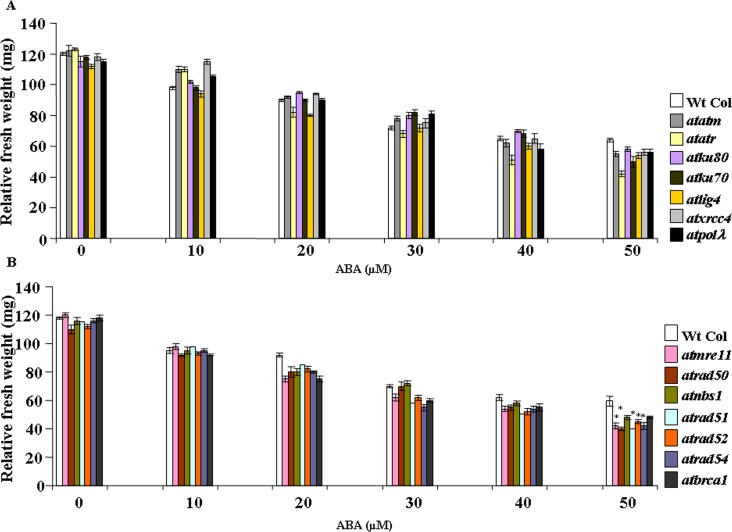
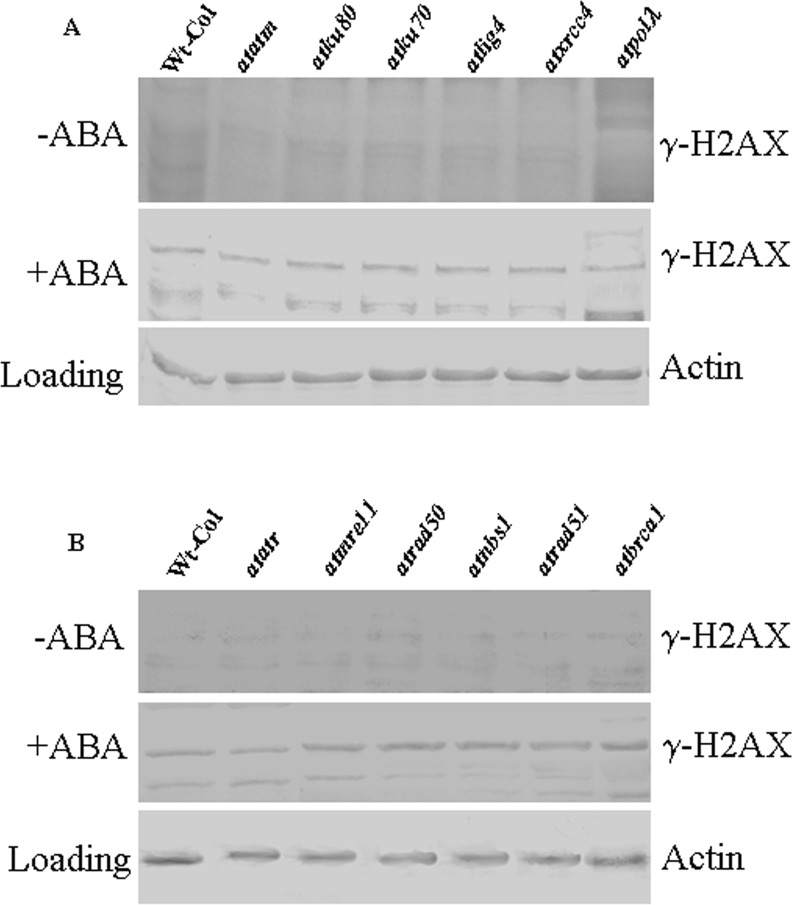
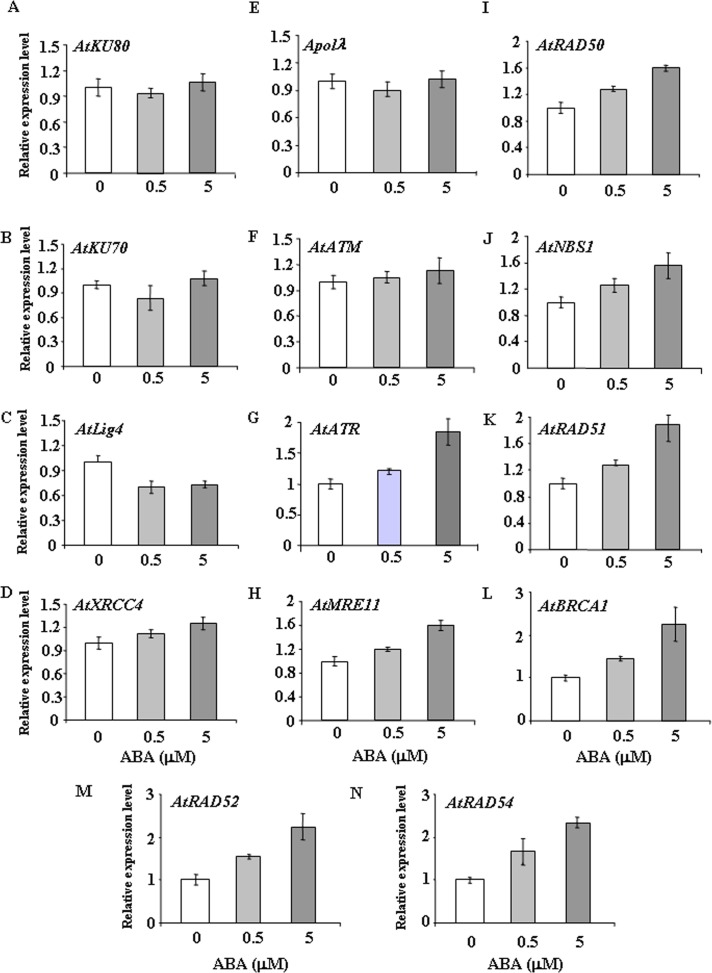
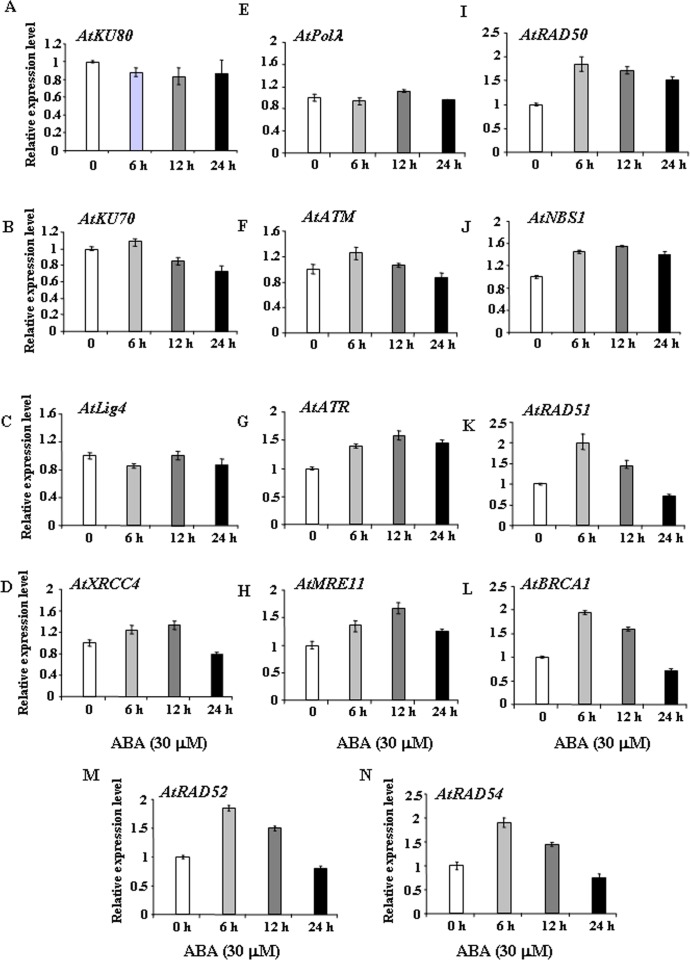
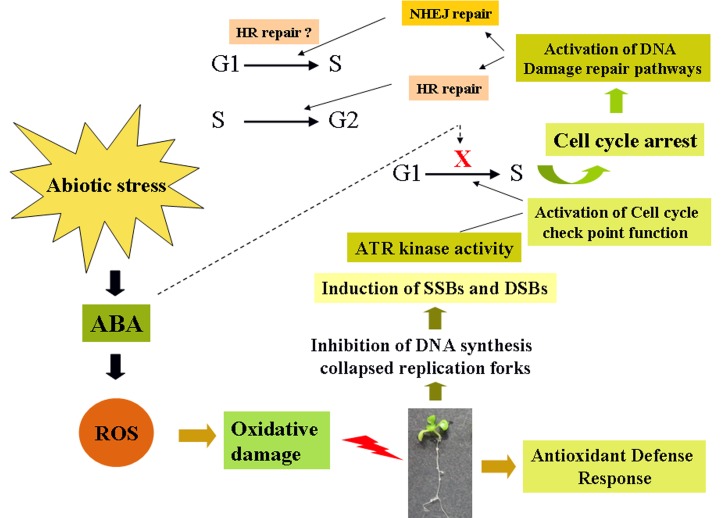
Abscisic acid (ABA) acts as an important plant hormone in regulating various aspects of plant growth and developmental processes particularly under abiotic stress conditions. An increased ABA level in plant cells inhibits DNA replication and cell division, causing plant growth retardation. In this study, we have investigated the effects of ABA on the growth responses of some major loss-of-function mutants of DNA double-stand break (DSB) repair genes in Arabidopsis during seed germination and early stages of seedling growth for understanding the role of ABA in the induction of genome instability in plants. A comparative analysis of ABA sensitivity of wild-type Arabidopsis and the knockout mutant lines related to DSB sensors, including atatm, atatr, the non-homologous end joining (NHEJ) pathway genes, and mutants related to homologous recombination (HR) pathway genes showed relatively enhanced sensitivity of atatr and HR-related mutants to ABA treatment. The expression levels of HR-related genes were increased in wild-type Arabidopsis (Col-0) during seed germination and early stages of seedling growth. Immunoblotting experiments detected phosphorylation of histone H2AX in wild-type (Col-0) and DSB repair gene mutants after ABA treatment, indicating the activation of DNA damage response due to ABA treatment. Analyses of DSB repair kinetics using comet assay under neutral condition have revealed comparatively slower DSB repair activity in HR mutants. Overall, our results have provided comprehensive information on the possible effect of ABA on DNA repair machinery in plants and also indicated potential functional involvement of HR pathway in repairing ABA induced DNA damage in Arabidopsis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
