

Review
doi: 10.1101/cshperspect.a028373.
The Role of Aneuploidy in Cancer Evolution
Affiliations
- PMID: 28049655
- PMCID: PMC5204330
- DOI: 10.1101/cshperspect.a028373
Item in Clipboard
Review
The Role of Aneuploidy in Cancer Evolution
Cold Spring Harb Perspect Med.
.
Abstract
Chromosomal aberrations during cell division represent one of the first recognized features of human cancer cells, and modern detection methods have revealed the pervasiveness of aneuploidy in cancer. The ongoing karyotypic changes brought about by chromosomal instability (CIN) contribute to tumor heterogeneity, drug resistance, and treatment failure. Whole-chromosome and segmental aneuploidies resulting from CIN have been proposed to allow "macroevolutionary" leaps that may contribute to profound phenotypic change. In this review, we will outline evidence indicating that aneuploidy and CIN contribute to cancer evolution.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
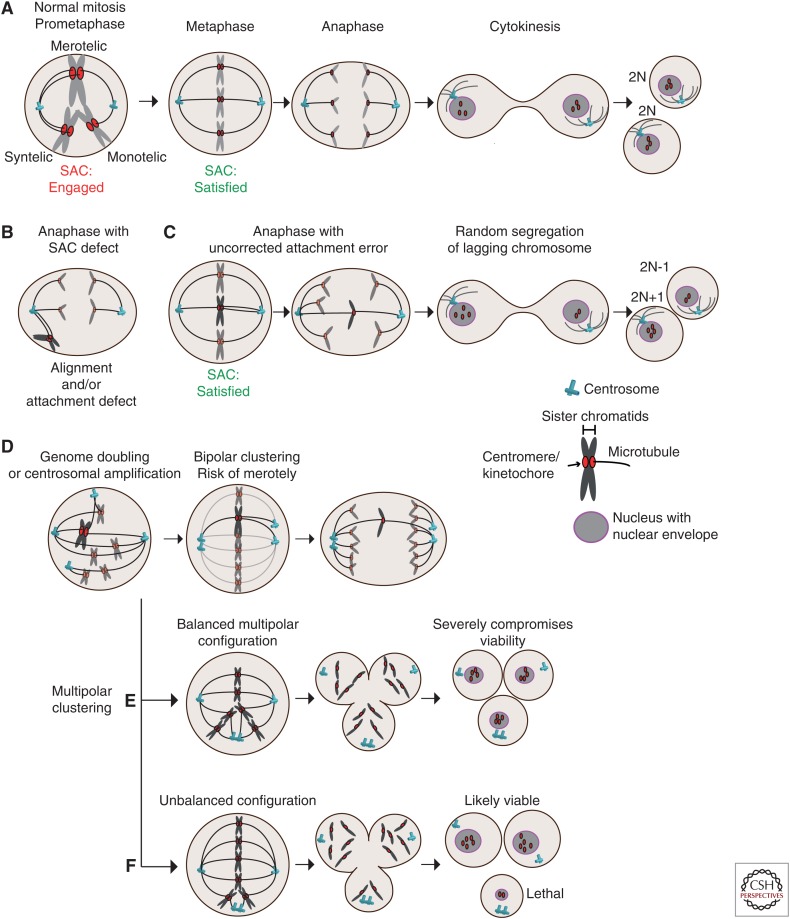
Mitotic defects leading to aneuploidy. (A) Kinetochore–microtubule attachment configurations and stages of mitosis. In early mitosis, improper attachments (syntelic and monotelic) activate the spindle assembly checkpoint (SAC) to delay anaphase onset. Merotelic misattachments poorly activate the checkpoint, but error correction mechanisms contribute to the formation of correct (amphiletic) bipolar attachments prior to chromosome separation at anaphase. (B) Cells with compromised SAC activity can enter anaphase with various attachment defects, here shown with syntelic chromosomes that did not congress to the metaphase plate. (C) Uncorrected merotelic attachments lead to lagging chromosomes at anaphase onset, causing random segregation, which can result in aneuploid daughter cells. (D) Cells with centrosome amplification and tetraploid cells enter mitosis with more than two centrosomes, creating a transient multipolar spindle. Bipolar clustering creates merotelic attachments because of the random clustering of the extra centrosomes. (E) Extra centrosome can result in a multipolar anaphase creating daughter cells lacking several chromosomes in a random fashion. (F) Random clustering can create a functional spindle pole that attaches only to few chromosomes, potentially resulting in viable aneuploidy in two out of three cells. Examples in D and E are especially well tolerated by tetraploid cells.

Mutations identified in genes important for chromosome segregation fidelity. Lollipop plots depicting the location and nature of mutations identified in 6807 tumors (32 cancer types) sequenced by The Cancer Genome Atlas (TCGA). Pathway analyses might shed new light on the frequency of chromosomal instability (CIN)-causing defects and the underlying mutations driving CIN, which will require a deeper understanding of the consequence of missense mutations. Many mutations occur in functional domains: for example, 33/67 (49%) of missense mutations in TTK (Mps1) occur in the kinase domain. Green dot, missense mutation; red dot, truncating mutation (nonsense, splice site, and frameshifts); purple dot, various mutation types. (Data and diagrams modified from cBioportal [Cerami et al. 2012; Gao et al. 2013].)
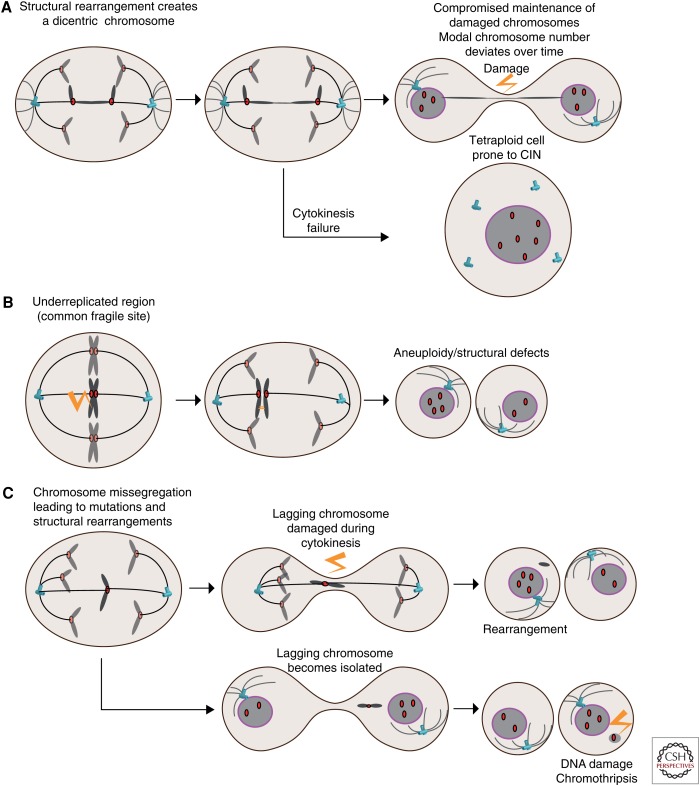
The interplay between whole-chromosome instability (wCIN), structural CIN (sCIN), and genomic instability (GIN). (A) Structural rearrangements caused by replication stress before mitosis created dicentric chromosomes. These can result in ultrafine bridges devoid of nuclear membrane and subject to extensive DNA damage and rearrangements akin to chromothripsis or hypermutation through kateagis and severing of the chromosome by exonucleases. Dicentric chromosomes can also prevent cytokinesis, as observed following telomere dysfunction. (B) Replication stress at common fragile sites triggers DNA replication during mitosis. Failing to complete DNA replication can lead to nondysjunction and aneuploidy. (C) Whole-chromosome missegregation can result in the trapping of the lagging chromosome during cytokinesis, causing DNA damage and structural aberrations. Alternatively, lagging chromosomes can become isolated in a micronucleus, in which the lack of nuclear membrane renders it susceptible to extensive DNA damage and chromothripsis. Damaged micronuclei can reintegrate the other chromosomes in the following division and be propagated.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources