

Cell Biology of the Caenorhabditis elegans Nucleus
- PMID: 28049702
- PMCID: PMC5216270
- DOI: 10.1534/genetics.116.197160
Cell Biology of the Caenorhabditis elegans Nucleus
Abstract
Studies on the Caenorhabditis elegans nucleus have provided fascinating insight to the organization and activities of eukaryotic cells. Being the organelle that holds the genetic blueprint of the cell, the nucleus is critical for basically every aspect of cell biology. The stereotypical development of C. elegans from a one cell-stage embryo to a fertile hermaphrodite with 959 somatic nuclei has allowed the identification of mutants with specific alterations in gene expression programs, nuclear morphology, or nuclear positioning. Moreover, the early C. elegans embryo is an excellent model to dissect the mitotic processes of nuclear disassembly and reformation with high spatiotemporal resolution. We review here several features of the C. elegans nucleus, including its composition, structure, and dynamics. We also discuss the spatial organization of chromatin and regulation of gene expression and how this depends on tight control of nucleocytoplasmic transport. Finally, the extensive connections of the nucleus with the cytoskeleton and their implications during development are described. Most processes of the C. elegans nucleus are evolutionarily conserved, highlighting the relevance of this powerful and versatile model organism to human biology.
Keywords: Caenorhabditis elegans; LEM-domain proteins; LINC; P granule; WormBook; chromatin; gene expression; lamin; nuclear envelope; nuclear pore complex; nucleocytoplasmic transport; nucleolus.
Copyright © 2017 by the Genetics Society of America.
Figures






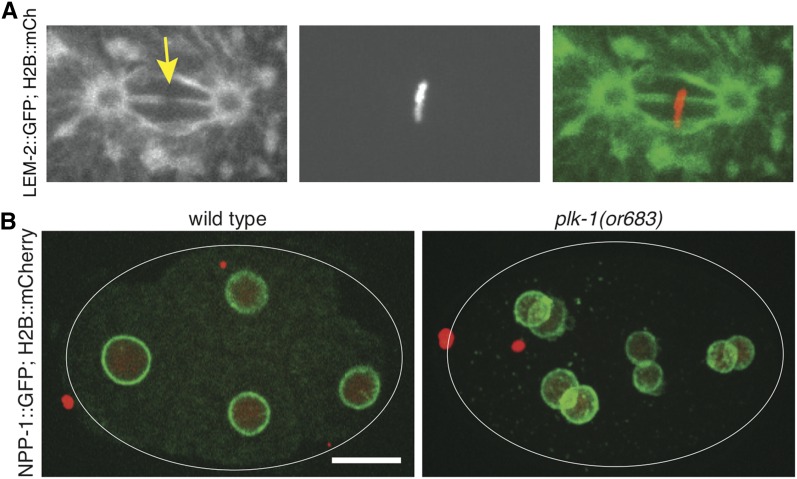









References
-
- Adam S. A., 2009. The nuclear transport machinery in Caenorhabditis elegans: a central role in morphogenesis. Semin. Cell Dev. Biol. 20: 576–581. - PubMed
-
- Anderson D. J., Hetzer M. W., 2007. Nuclear envelope formation by chromatin-mediated reorganization of the endoplasmic reticulum. Nat. Cell Biol. 9: 1160–1166. - PubMed
-
- Asencio C., Davidson I. F., Santarella-Mellwig R., Ly-Hartig T. B., Mall M., et al. , 2012. Coordination of kinase and phosphatase activities by Lem4 enables nuclear envelope reassembly during mitosis. Cell 150: 122–135. - PubMed
-
- Askjaer P., Ercan S., Meister P., 2014a Modern techniques for the analysis of chromatin and nuclear organization in C. elegans (April 2, 2014), WormBook, ed. The C. elegans Research Community WormBook, 10.1895/wormbook.1.169.1, http://wormbook.org. - PMC - PubMed
-
- Askjaer P., Galy V., Meister P., 2014b Modern tools to study nuclear pore complexes and nucleocytoplasmic transport in Caenorhabditis elegans. Methods Cell Biol. 122: 277–310. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
