

Essential Role for Polycomb Group Protein Pcgf6 in Embryonic Stem Cell Maintenance and a Noncanonical Polycomb Repressive Complex 1 (PRC1) Integrity
- PMID: 28049731
- PMCID: PMC5314173
- DOI: 10.1074/jbc.M116.763961
Essential Role for Polycomb Group Protein Pcgf6 in Embryonic Stem Cell Maintenance and a Noncanonical Polycomb Repressive Complex 1 (PRC1) Integrity
Abstract
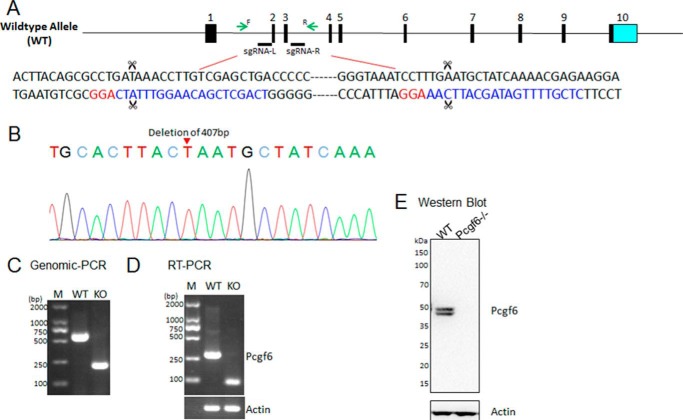
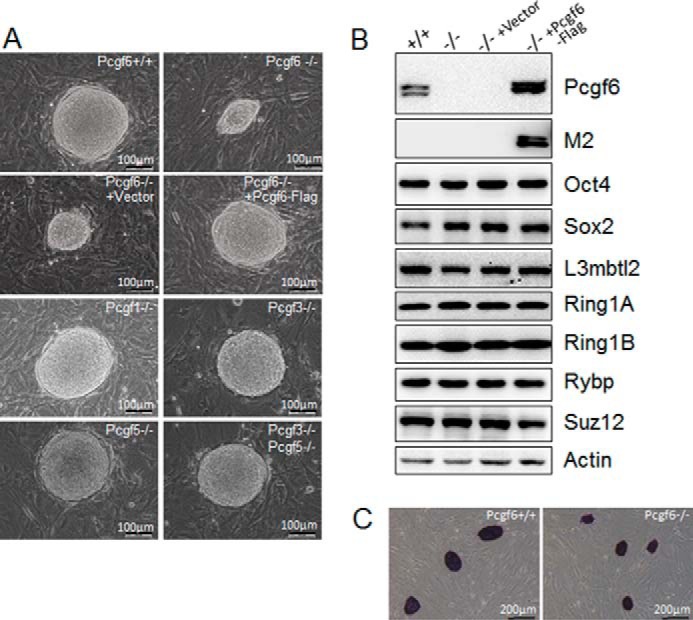
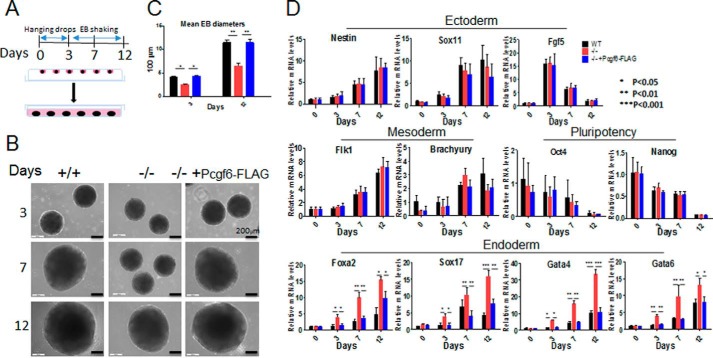
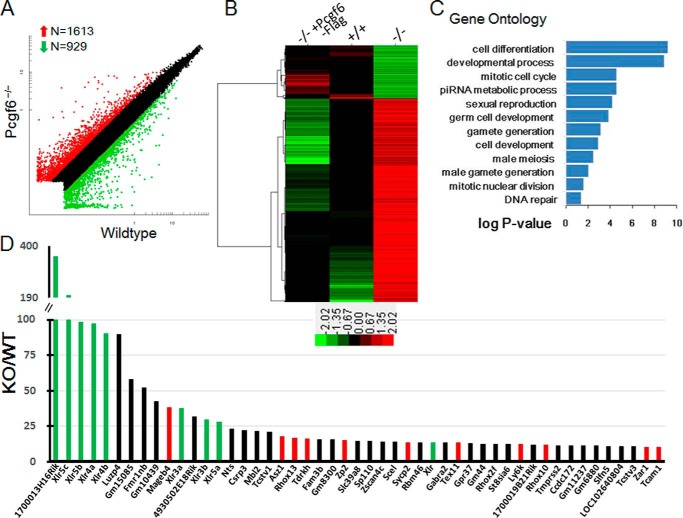
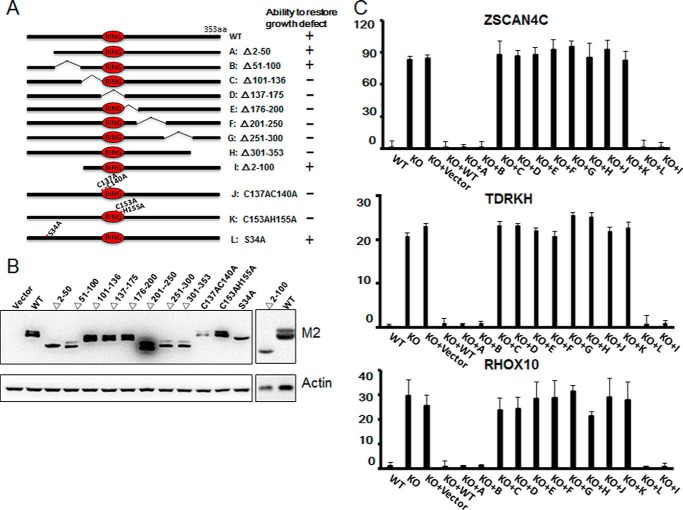
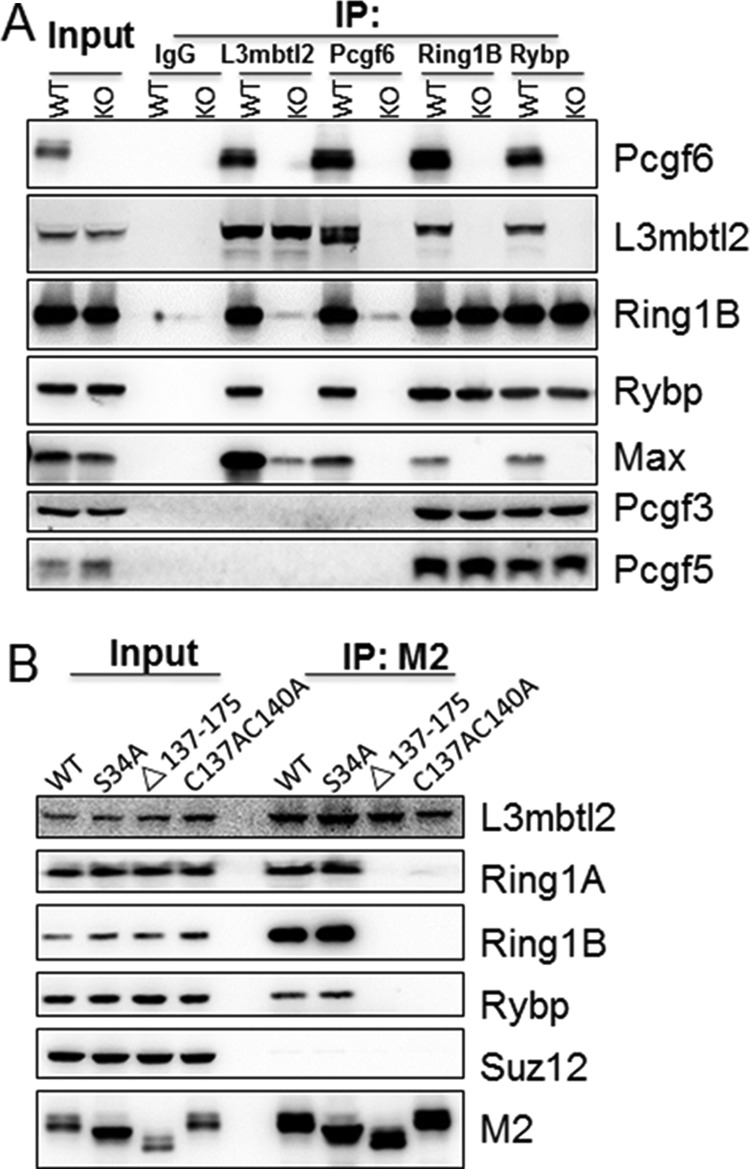
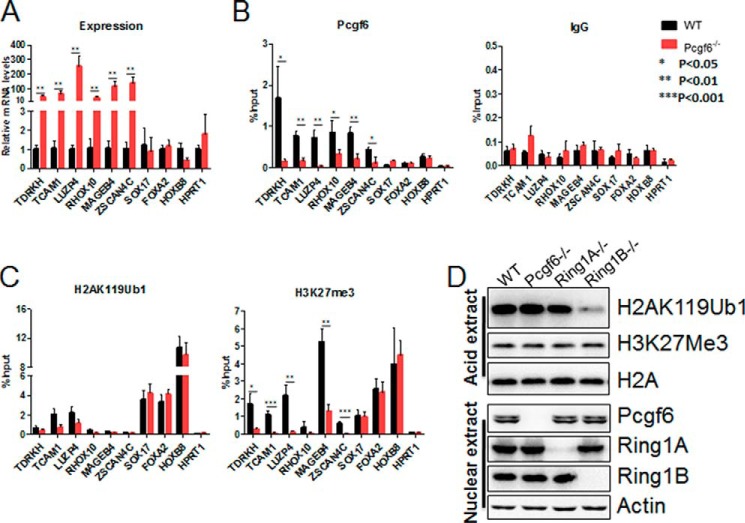
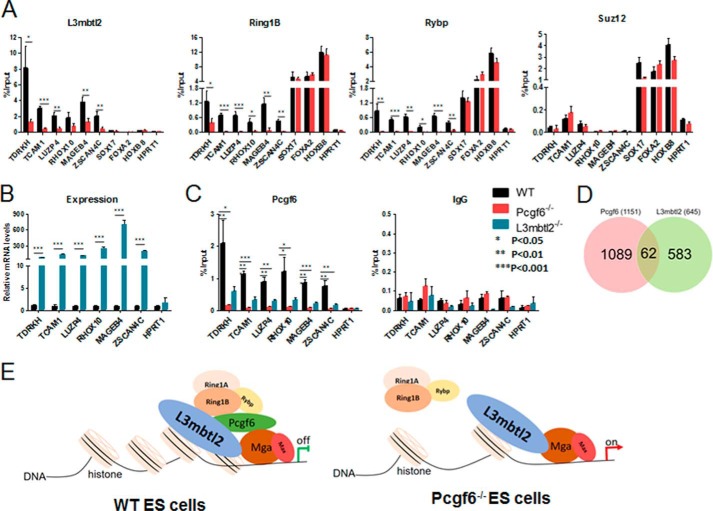
The Polycomb group (PcG) proteins have an important role in controlling the expression of key genes implicated in embryonic development, differentiation, and decision of cell fates. Emerging evidence suggests that Polycomb repressive complexes 1 (PRC1) is defined by the six Polycomb group RING finger protein (Pcgf) paralogs, and Pcgf proteins can assemble into noncanonical PRC1 complexes. However, little is known about the precise mechanisms of differently composed noncanonical PRC1 in the maintenance of the pluripotent cell state. Here we disrupt the Pcgf genes in mouse embryonic stem cells by CRISPR-Cas9 and find Pcgf6 null embryonic stem cells display severe defects in self-renewal and differentiation. Furthermore, Pcgf6 regulates genes mostly involved in differentiation and spermatogenesis by assembling a noncanonical PRC1 complex PRC1.6. Notably, Pcgf6 deletion causes a dramatic decrease in PRC1.6 binding to target genes and no loss of H2AK119ub1. Thus, Pcgf6 is essential for recruitment of PRC1.6 to chromatin. Our results reveal a previously uncharacterized, H2AK119ub1-independent chromatin assembly associated with PRC1.6 complex.
Keywords: Polycomb; chromatin; embryonic stem cell; epigenetics; transcription regulation.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Pcgf6, a polycomb group protein, regulates mesodermal lineage differentiation in murine ESCs and functions in iPS reprogramming.Stem Cells. 2014 Dec;32(12):3112-25. doi: 10.1002/stem.1826. Stem Cells. 2014. PMID: 25187489
-
Functional redundancy among Polycomb complexes in maintaining the pluripotent state of embryonic stem cells.Stem Cell Reports. 2022 May 10;17(5):1198-1214. doi: 10.1016/j.stemcr.2022.02.020. Epub 2022 Mar 31. Stem Cell Reports. 2022. PMID: 35364009 Free PMC article.
-
PCGF6-PRC1 suppresses premature differentiation of mouse embryonic stem cells by regulating germ cell-related genes.Elife. 2017 Mar 17;6:e21064. doi: 10.7554/eLife.21064. Elife. 2017. PMID: 28304275 Free PMC article.
-
[Controlling the spatiotemporal expression of germ line specific genes by PRC1.6 complex].Yi Chuan. 2019 Apr 20;41(4):271-284. doi: 10.16288/j.yczz.18-332. Yi Chuan. 2019. PMID: 30992249 Review. Chinese.
-
Polycomb group proteins and their roles in regulating stem cell development.Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2012 Jun;34(3):281-5. doi: 10.3881/j.issn.1000-503X.2012.03.019. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2012. PMID: 22776663 Review.
Cited by
-
Polycomb group ring finger protein 6 suppresses Myc-induced lymphomagenesis.Life Sci Alliance. 2022 Apr 14;5(8):e202101344. doi: 10.26508/lsa.202101344. Print 2022 Aug. Life Sci Alliance. 2022. PMID: 35422437 Free PMC article.
-
Synergy between Variant PRC1 Complexes Defines Polycomb-Mediated Gene Repression.Mol Cell. 2019 Jun 6;74(5):1020-1036.e8. doi: 10.1016/j.molcel.2019.03.024. Epub 2019 Apr 24. Mol Cell. 2019. PMID: 31029541 Free PMC article.
-
Polycomb Assemblies Multitask to Regulate Transcription.Epigenomes. 2019 Jun 20;3(2):12. doi: 10.3390/epigenomes3020012. Epigenomes. 2019. PMID: 34968234 Free PMC article. Review.
-
Repression of germline genes by PRC1.6 and SETDB1 in the early embryo precedes DNA methylation-mediated silencing.Nat Commun. 2021 Dec 2;12(1):7020. doi: 10.1038/s41467-021-27345-x. Nat Commun. 2021. PMID: 34857746 Free PMC article.
-
A molecular roadmap for the emergence of early-embryonic-like cells in culture.Nat Genet. 2018 Jan;50(1):106-119. doi: 10.1038/s41588-017-0016-5. Epub 2017 Dec 18. Nat Genet. 2018. PMID: 29255263 Free PMC article.
References
-
- Simon J. A., and Kingston R. E. (2009) Mechanisms of polycomb gene silencing: knowns and unknowns. Nat. Rev. Mol. Cell Biol. 10, 697–708 - PubMed
-
- Piunti A., and Shilatifard A. (2016) Epigenetic balance of gene expression by Polycomb and COMPASS families. Science 352, aad9780. - PubMed
-
- Di Croce L., and Helin K. (2013) Transcriptional regulation by Polycomb group proteins. Nat. Struct. Mol. Biol. 20, 1147–1155 - PubMed
-
- Tavares L., Dimitrova E., Oxley D., Webster J., Poot R., Demmers J., Bezstarosti K., Taylor S., Ura H., Koide H., Wutz A., Vidal M., Elderkin S., and Brockdorff N. (2012) RYBP-PRC1 complexes mediate H2A ubiquitylation at polycomb target sites independently of PRC2 and H3K27me3. Cell 148, 664–678 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
