

Outside-Xylem Vulnerability, Not Xylem Embolism, Controls Leaf Hydraulic Decline during Dehydration
- PMID: 28049739
- PMCID: PMC5291720
- DOI: 10.1104/pp.16.01643
Outside-Xylem Vulnerability, Not Xylem Embolism, Controls Leaf Hydraulic Decline during Dehydration
Abstract
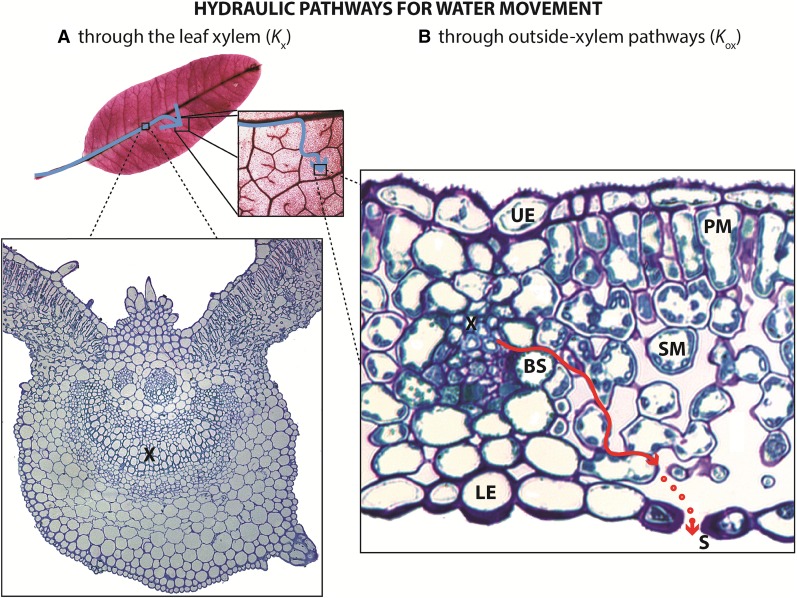
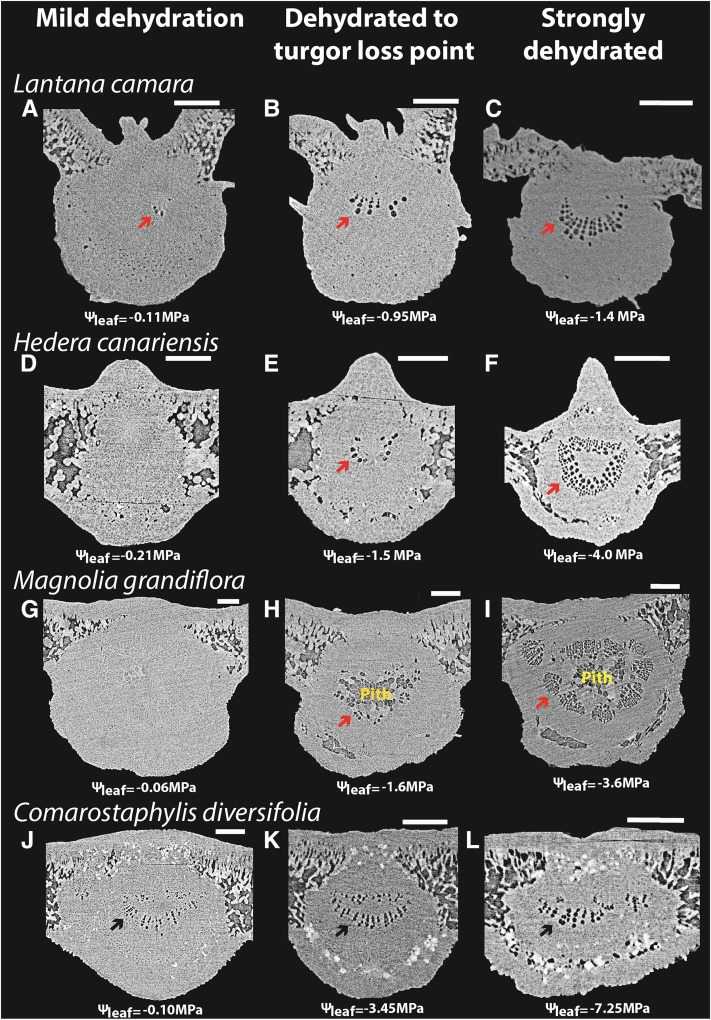
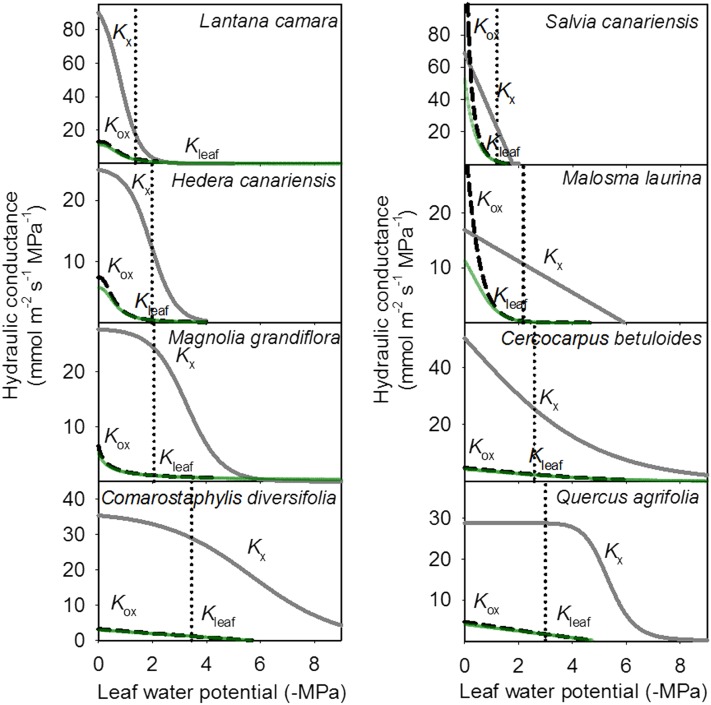
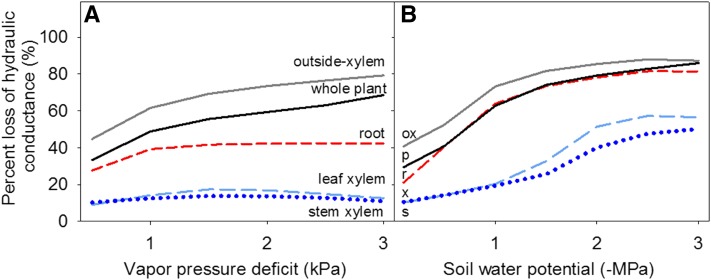
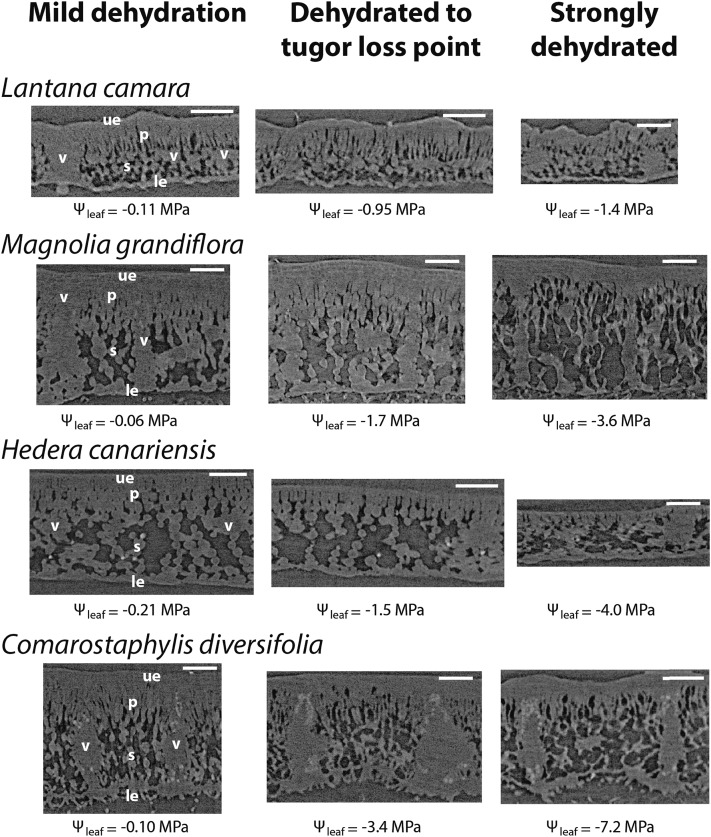
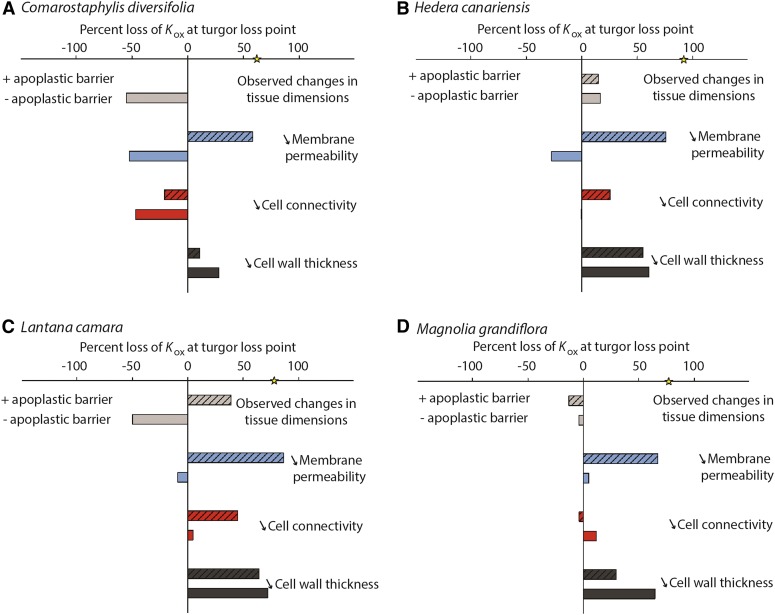
Leaf hydraulic supply is crucial to maintaining open stomata for CO2 capture and plant growth. During drought-induced dehydration, the leaf hydraulic conductance (Kleaf) declines, which contributes to stomatal closure and, eventually, to leaf death. Previous studies have tended to attribute the decline of Kleaf to embolism in the leaf vein xylem. We visualized at high resolution and quantified experimentally the hydraulic vulnerability of xylem and outside-xylem pathways and modeled their respective influences on plant water transport. Evidence from all approaches indicated that the decline of Kleaf during dehydration arose first and foremost due to the vulnerability of outside-xylem tissues. In vivo x-ray microcomputed tomography of dehydrating leaves of four diverse angiosperm species showed that, at the turgor loss point, only small fractions of leaf vein xylem conduits were embolized, and substantial xylem embolism arose only under severe dehydration. Experiments on an expanded set of eight angiosperm species showed that outside-xylem hydraulic vulnerability explained 75% to 100% of Kleaf decline across the range of dehydration from mild water stress to beyond turgor loss point. Spatially explicit modeling of leaf water transport pointed to a role for reduced membrane conductivity consistent with published data for cells and tissues. Plant-scale modeling suggested that outside-xylem hydraulic vulnerability can protect the xylem from tensions that would induce embolism and disruption of water transport under mild to moderate soil and atmospheric droughts. These findings pinpoint outside-xylem tissues as a central locus for the control of leaf and plant water transport during progressive drought.
© 2017 The author(s). All Rights Reserved.
Figures
References
-
- Anderegg WRL, Flint A, Huang C, Flint L, Berry JA, Davis FW, Sperry JS, Field CB (2015) Tree mortality predicted from drought-induced vascular damage. Nat Geosci 8: 367–371
-
- Bartlett MK, Scoffoni C, Sack L (2012) The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis. Ecol Lett 15: 393–405 - PubMed
-
- Blackman CJ, Brodribb TJ, Jordan GJ (2010) Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol 188: 1113–1123 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
