

Tbx20 controls the expression of the KCNH2 gene and of hERG channels
- PMID: 28049825
- PMCID: PMC5255604
- DOI: 10.1073/pnas.1612383114
Tbx20 controls the expression of the KCNH2 gene and of hERG channels
Abstract
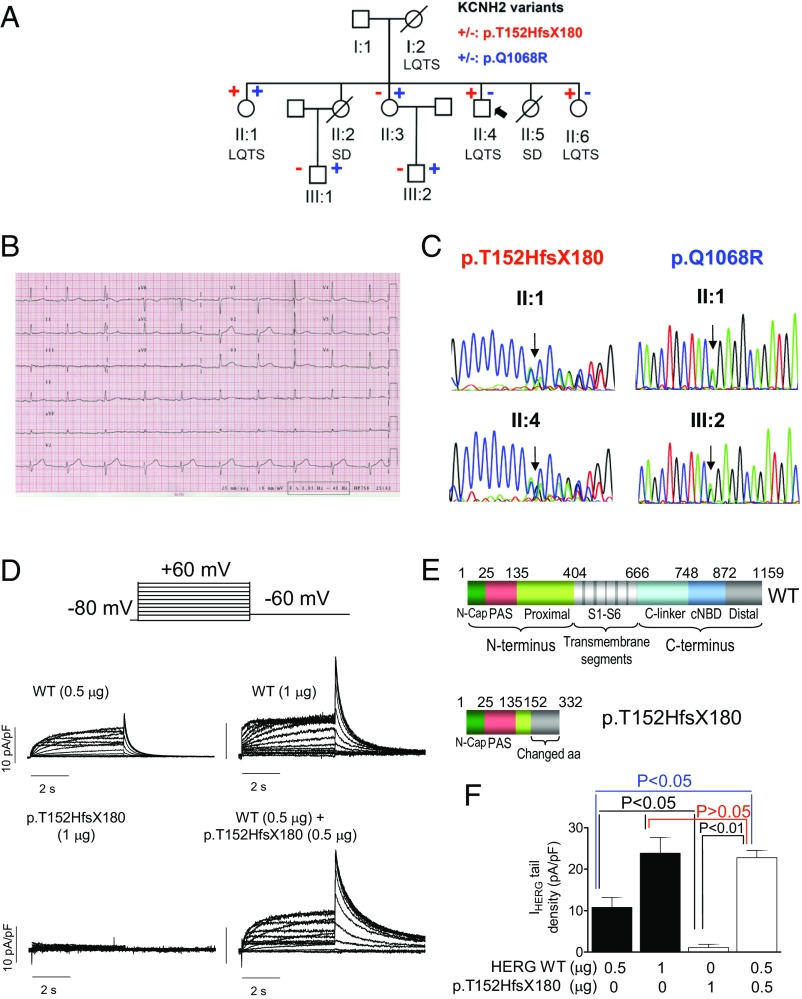
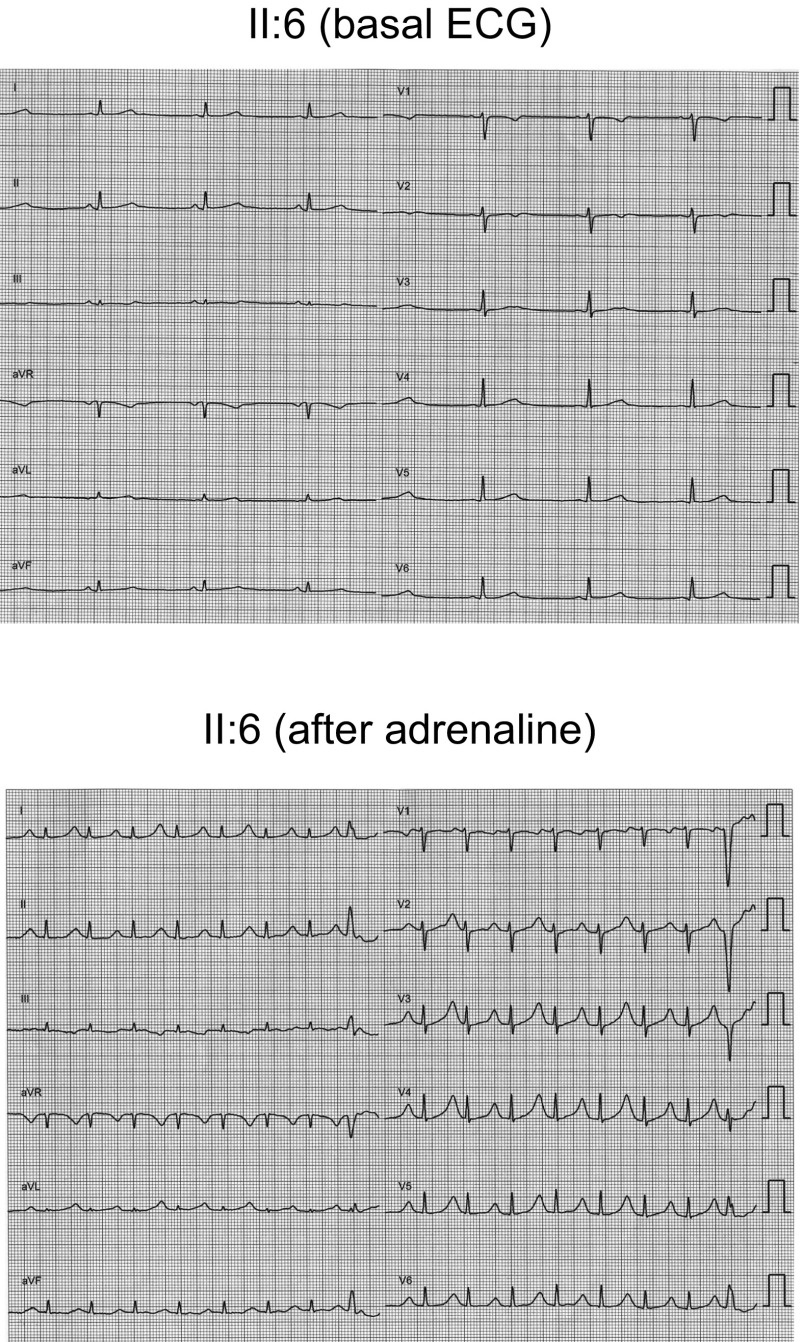
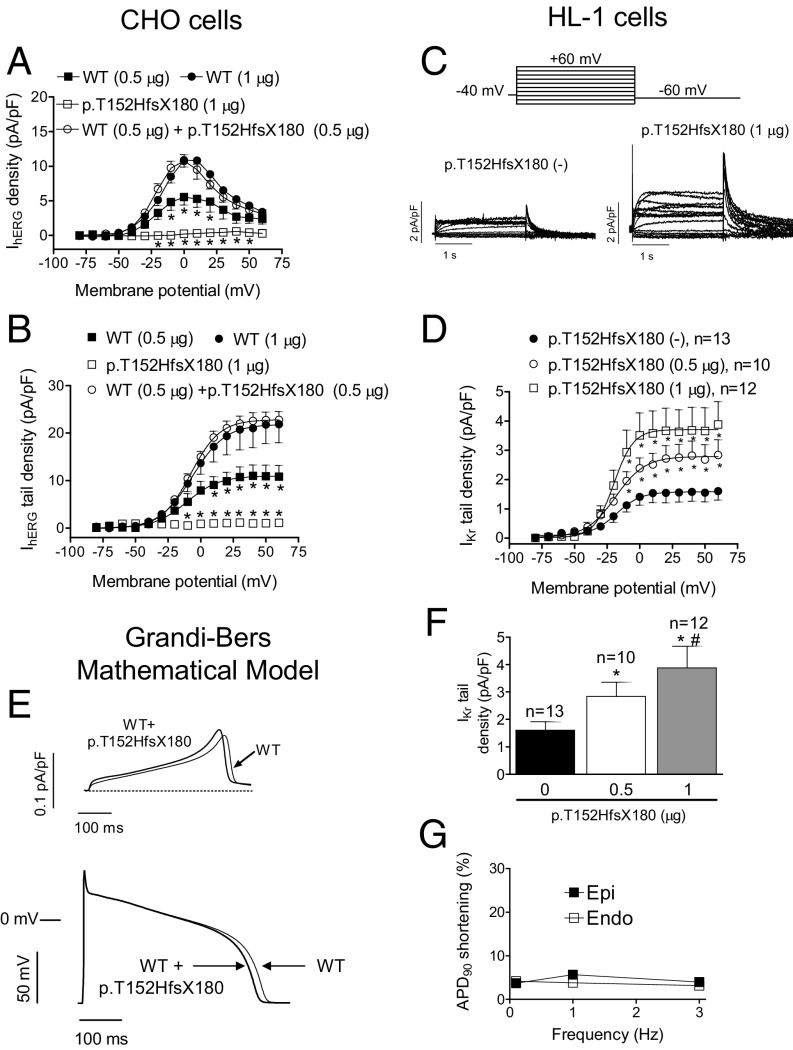
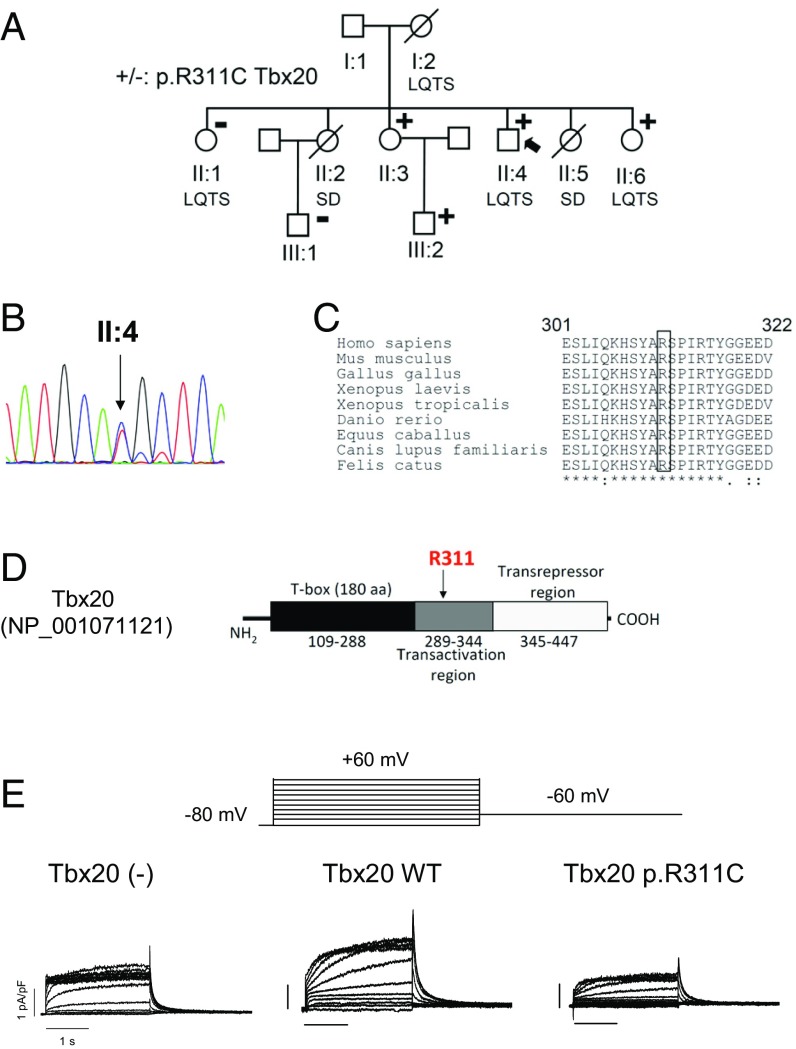
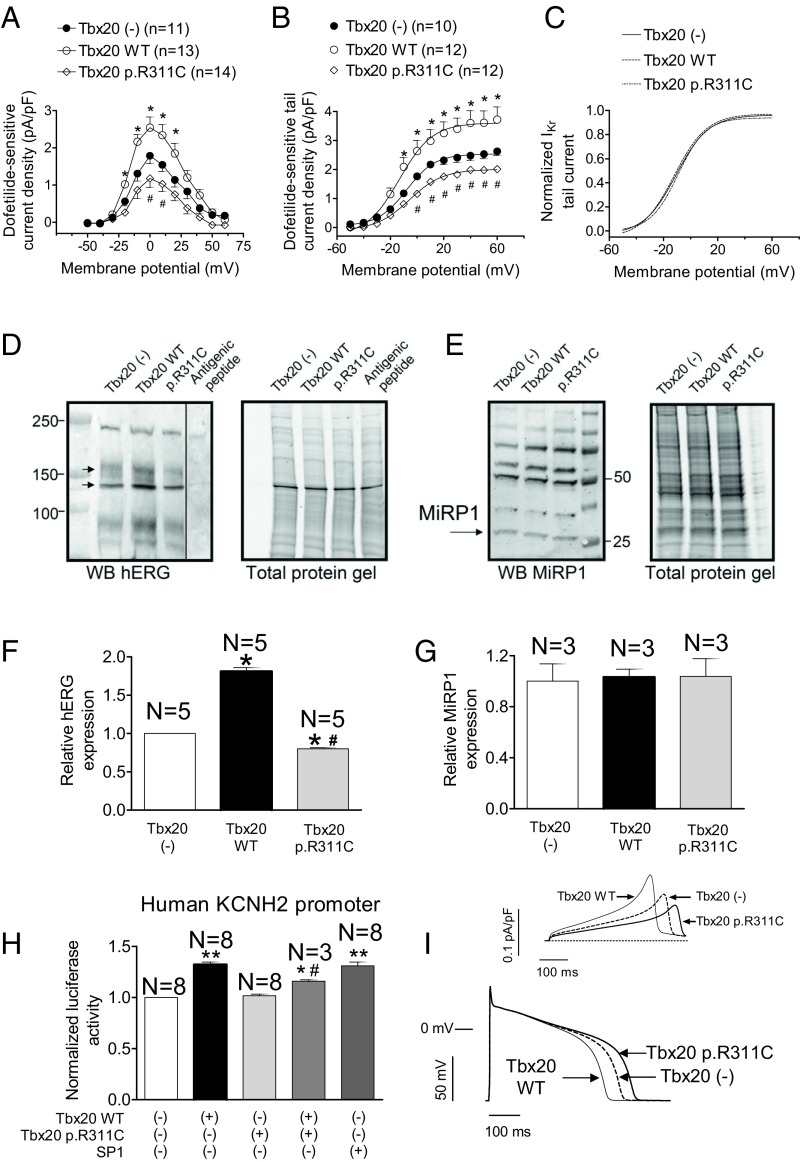
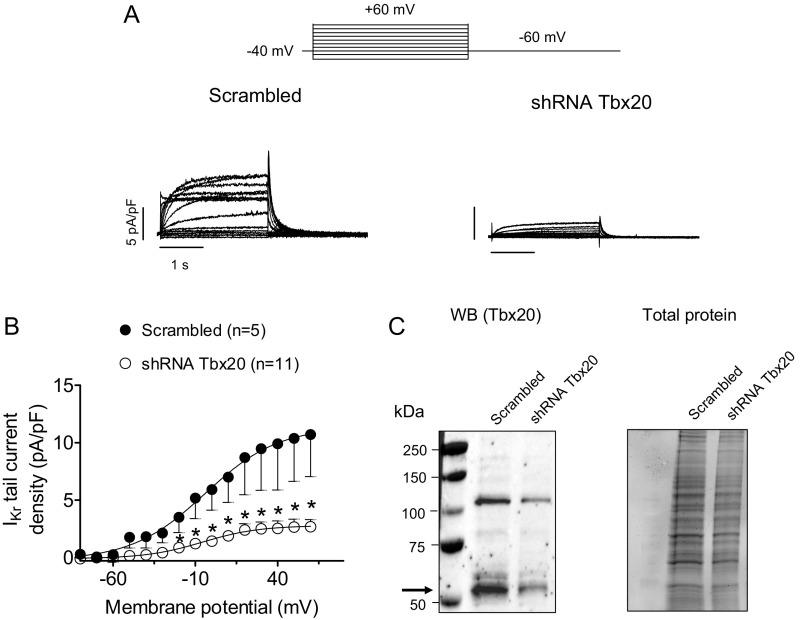
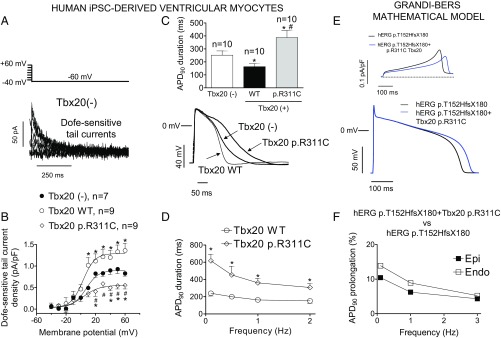
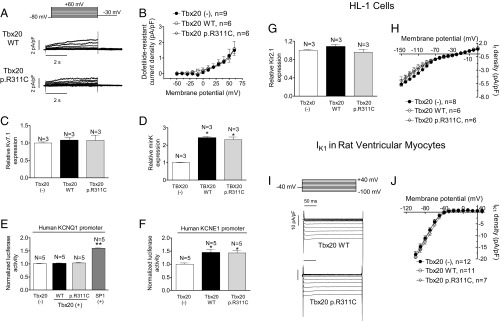
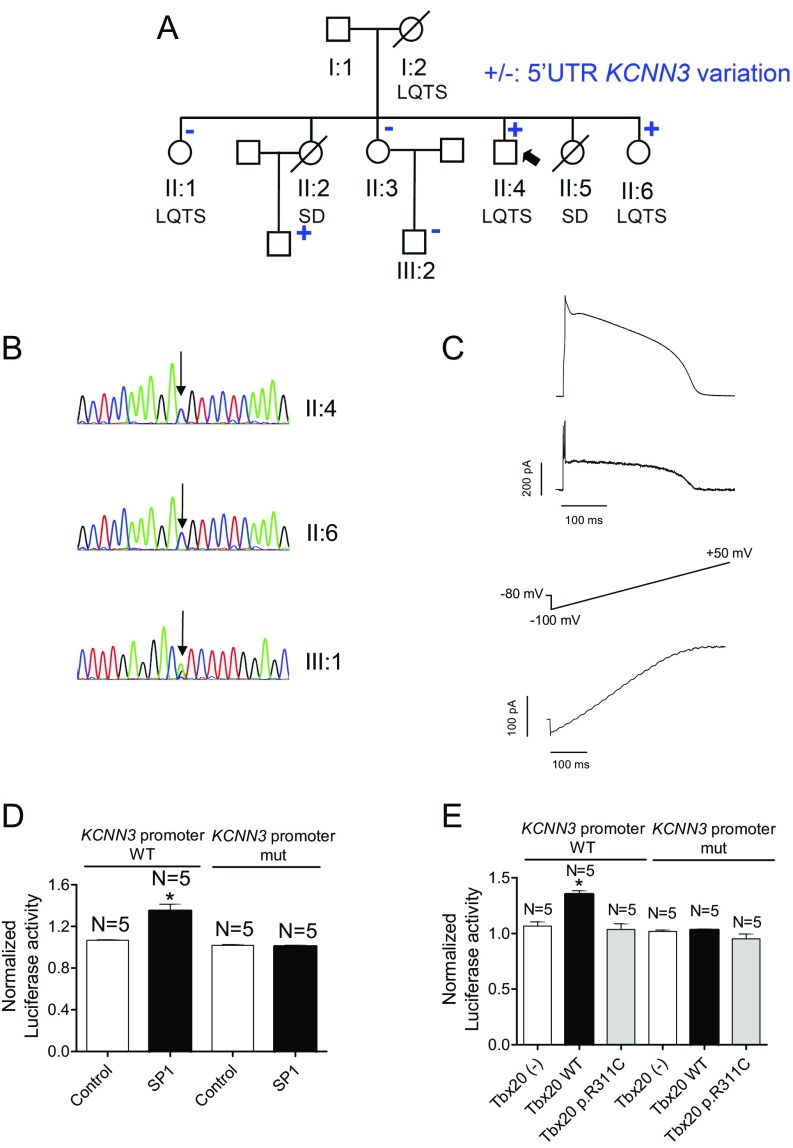
Long QT syndrome (LQTS) exhibits great phenotype variability among family members carrying the same mutation, which can be partially attributed to genetic factors. We functionally analyzed the KCNH2 (encoding for Kv11.1 or hERG channels) and TBX20 (encoding for the transcription factor Tbx20) variants found by next-generation sequencing in two siblings with LQTS in a Spanish family of African ancestry. Affected relatives harbor a heterozygous mutation in KCNH2 that encodes for p.T152HfsX180 Kv11.1 (hERG). This peptide, by itself, failed to generate any current when transfected into Chinese hamster ovary (CHO) cells but, surprisingly, exerted "chaperone-like" effects over native hERG channels in both CHO cells and mouse atrial-derived HL-1 cells. Therefore, heterozygous transfection of native (WT) and p.T152HfsX180 hERG channels generated a current that was indistinguishable from that generated by WT channels alone. Some affected relatives also harbor the p.R311C mutation in Tbx20. In human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs), Tbx20 enhanced human KCNH2 gene expression and hERG currents (IhERG) and shortened action-potential duration (APD). However, Tbx20 did not modify the expression or activity of any other channel involved in ventricular repolarization. Conversely, p.R311C Tbx20 did not increase KCNH2 expression in hiPSC-CMs, which led to decreased IhERG and increased APD. Our results suggest that Tbx20 controls the expression of hERG channels responsible for the rapid component of the delayed rectifier current. On the contrary, p.R311C Tbx20 specifically disables the Tbx20 protranscriptional activity over KCNH2 Therefore, TBX20 can be considered a KCNH2-modifying gene.
Keywords: Tbx20; cardiomyocytes; hERG channels; human induced pluripotent stem cells; long QT syndrome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
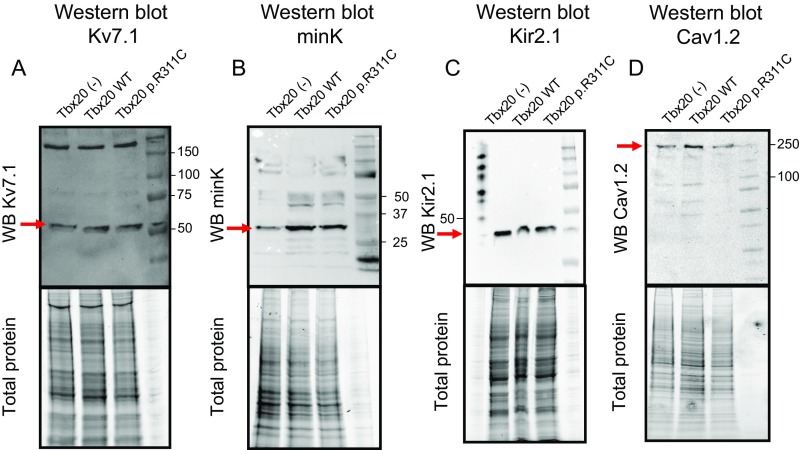
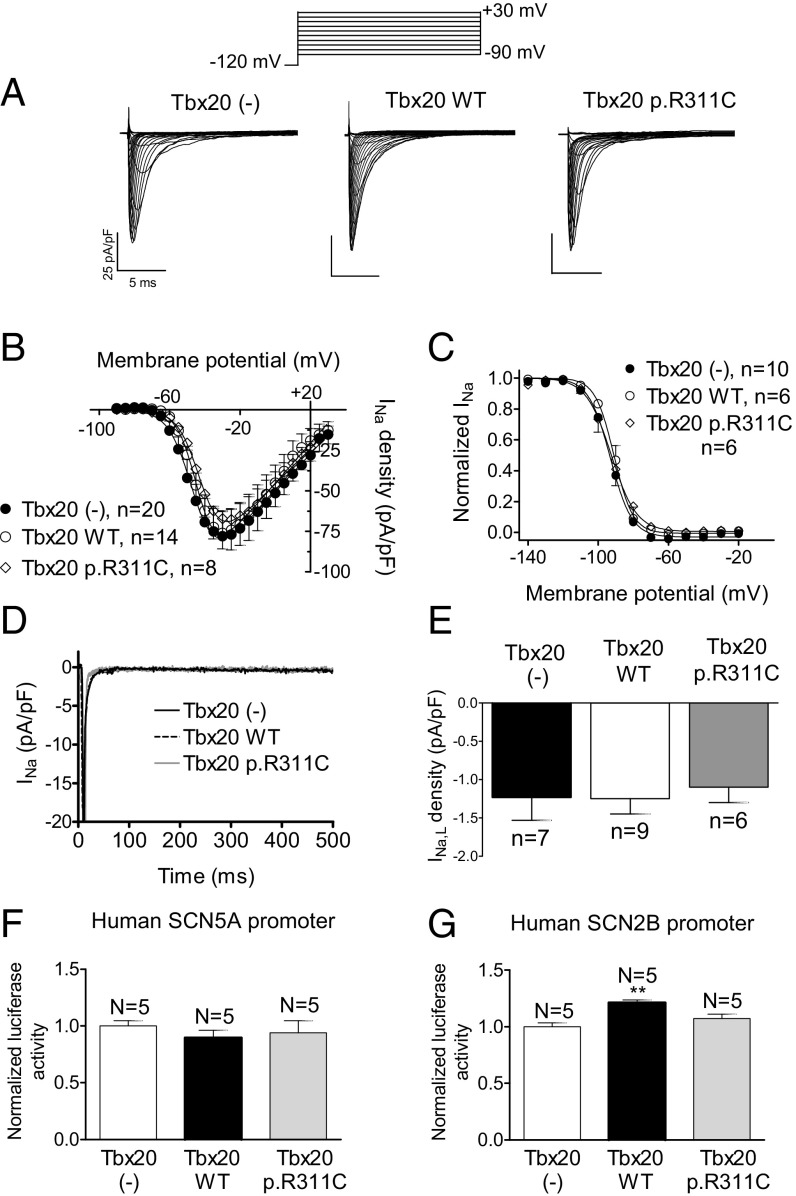
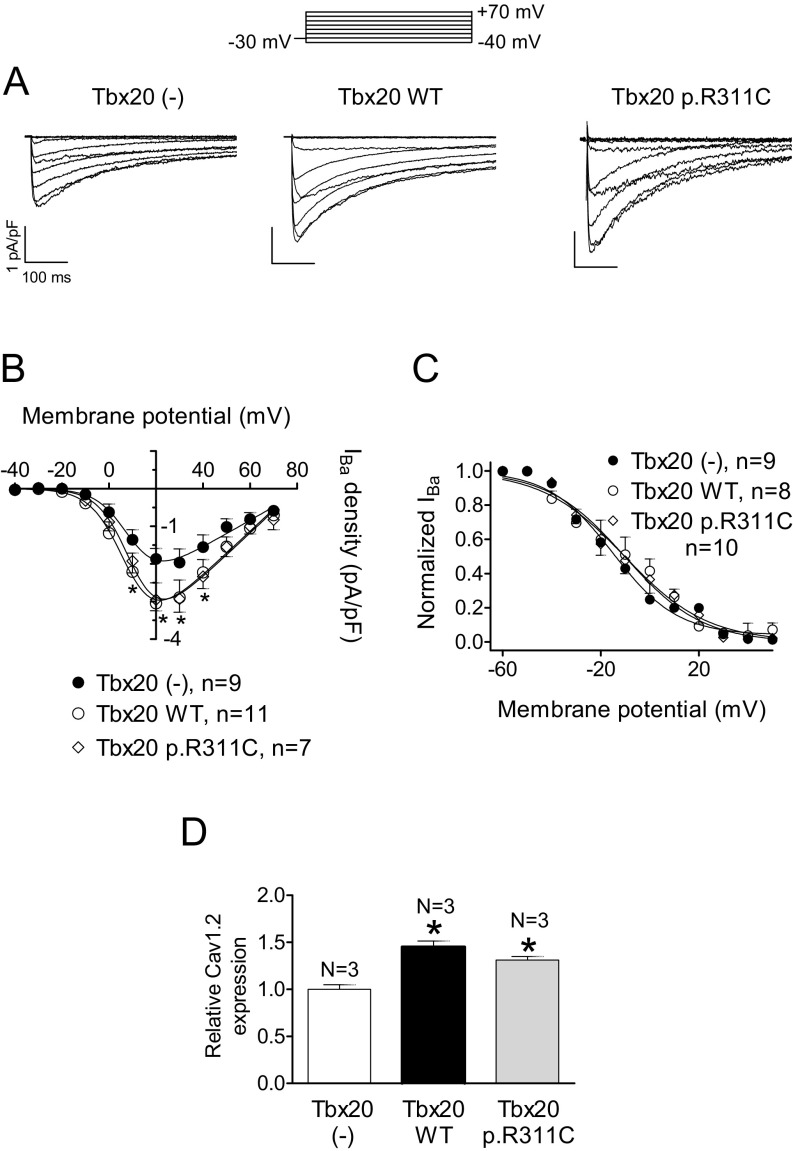
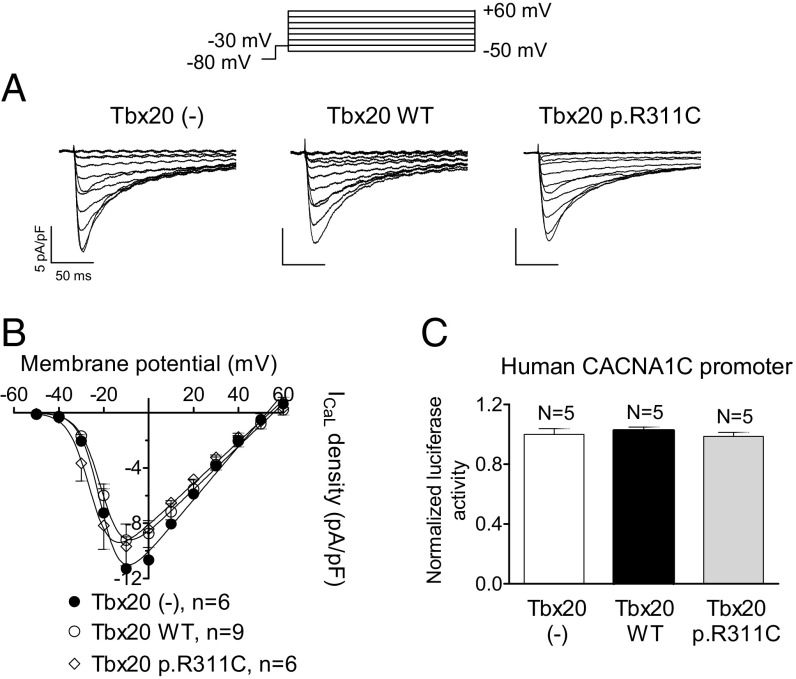
References
-
- Nakano Y, Shimizu W. Genetics of long-QT syndrome. J Hum Genet. 2016;61(1):51–55. - PubMed
-
- Havakuk O, Viskin S. A tale of 2 diseases: The history of long-QT syndrome and Brugada syndrome. J Am Coll Cardiol. 2016;67(1):100–108. - PubMed
-
- Keating MT, Sanguinetti MC. Molecular and cellular mechanisms of cardiac arrhythmias. Cell. 2001;104(4):569–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
