

Characterization of cytopathic factors through genome-wide analysis of the Zika viral proteins in fission yeast
- PMID: 28049830
- PMCID: PMC5255628
- DOI: 10.1073/pnas.1619735114
Characterization of cytopathic factors through genome-wide analysis of the Zika viral proteins in fission yeast
Abstract
The Zika virus (ZIKV) causes microcephaly and the Guillain-Barré syndrome. Little is known about how ZIKV causes these conditions or which ZIKV viral protein(s) is responsible for the associated ZIKV-induced cytopathic effects, including cell hypertrophy, growth restriction, cell-cycle dysregulation, and cell death. We used fission yeast for the rapid, global functional analysis of the ZIKV genome. All 14 proteins or small peptides were produced under an inducible promoter, and we measured the intracellular localization and the specific effects on ZIKV-associated cytopathic activities of each protein. The subcellular localization of each ZIKV protein was in overall agreement with its predicted protein structure. Five structural and two nonstructural ZIKV proteins showed various levels of cytopathic effects. The expression of these ZIKV proteins restricted cell proliferation, induced hypertrophy, or triggered cellular oxidative stress leading to cell death. The expression of premembrane protein (prM) resulted in cell-cycle G1 accumulation, whereas membrane-anchored capsid (anaC), membrane protein (M), envelope protein (E), and nonstructural protein 4A (NS4A) caused cell-cycle G2/M accumulation. A mechanistic study revealed that NS4A-induced cellular hypertrophy and growth restriction were mediated specifically through the target of rapamycin (TOR) cellular stress pathway involving Tor1 and type 2A phosphatase activator Tip41. These findings should provide a reference for future research on the prevention and treatment of ZIKV diseases.
Keywords: Schizosaccharomyces pombe; Zika genome; Zika proteins; cytopathic factors; fission yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

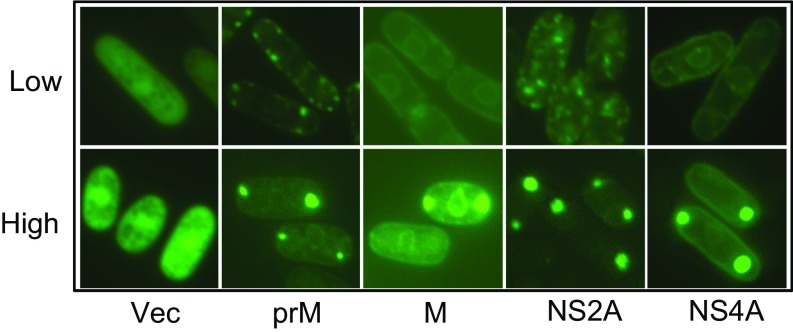



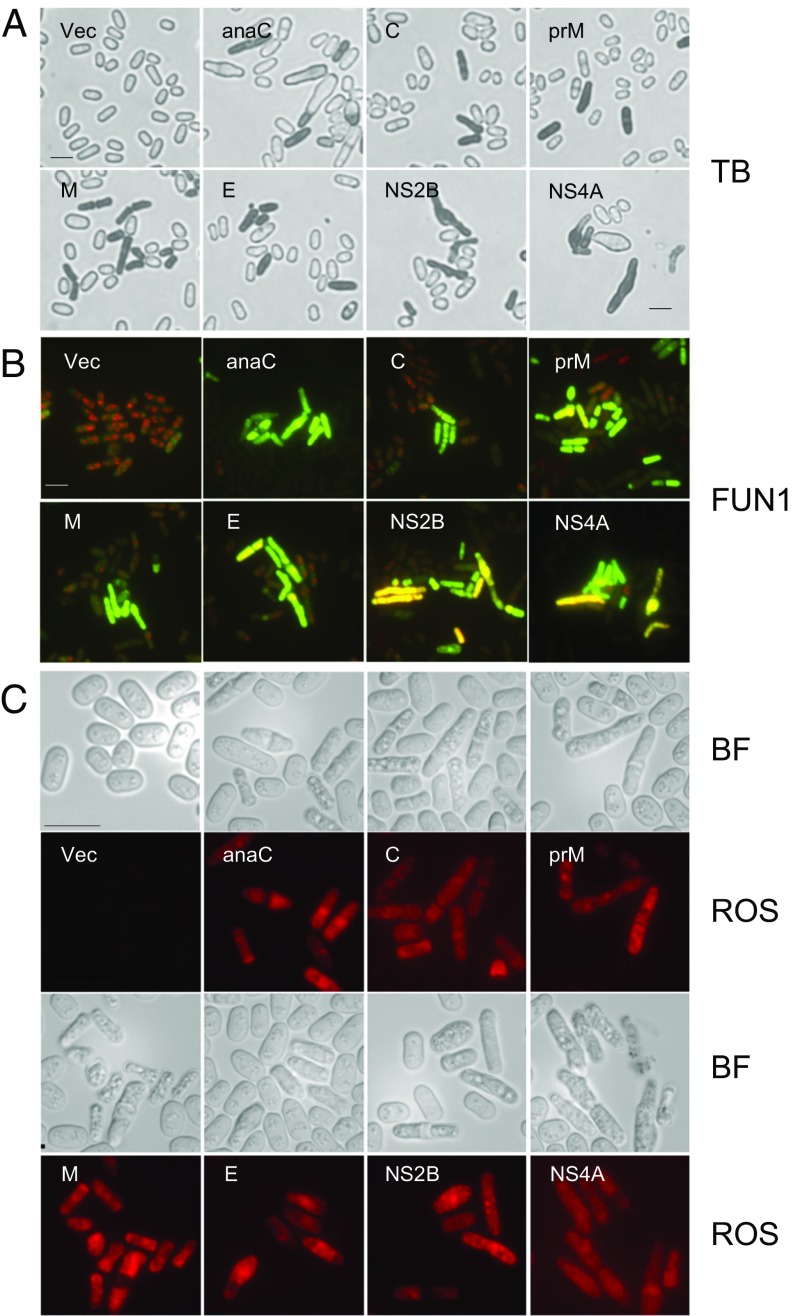

References
-
- Smith DW, Mackenzie J. Zika virus and Guillain-Barré syndrome: Another viral cause to add to the list. Lancet. 2016;387(10027):1486–1488. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
