

Co-Option and De Novo Gene Evolution Underlie Molluscan Shell Diversity
- PMID: 28053006
- PMCID: PMC5400390
- DOI: 10.1093/molbev/msw294
Co-Option and De Novo Gene Evolution Underlie Molluscan Shell Diversity
Abstract
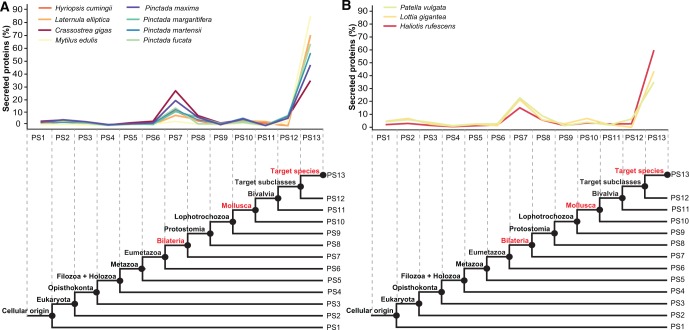
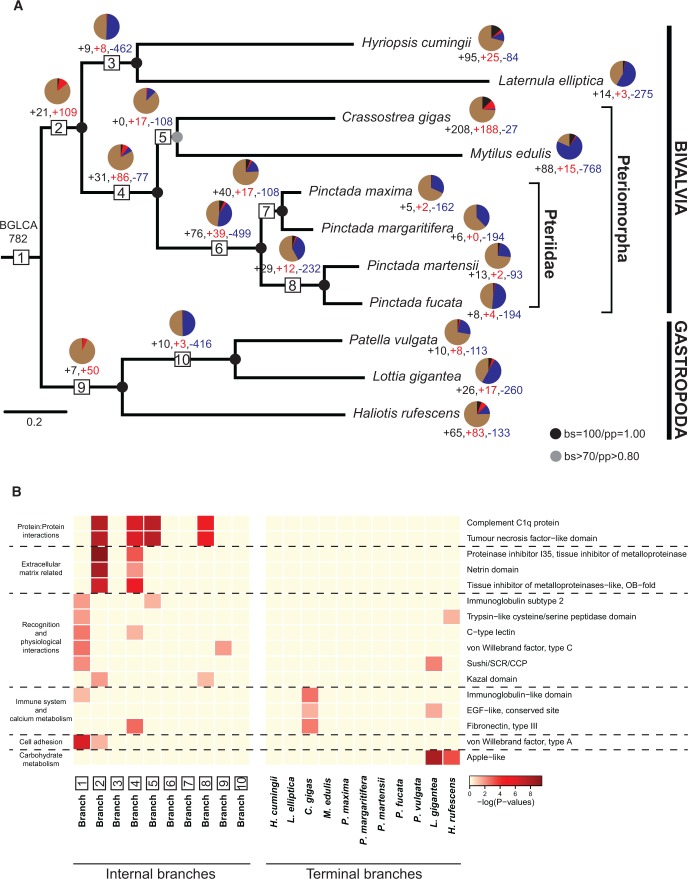
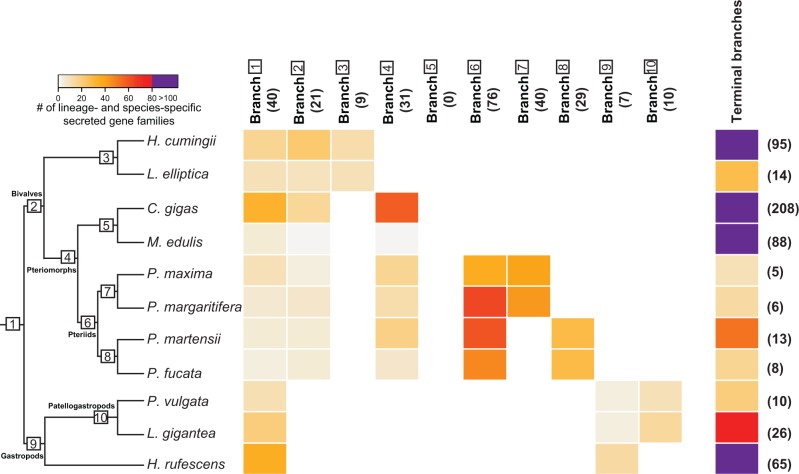
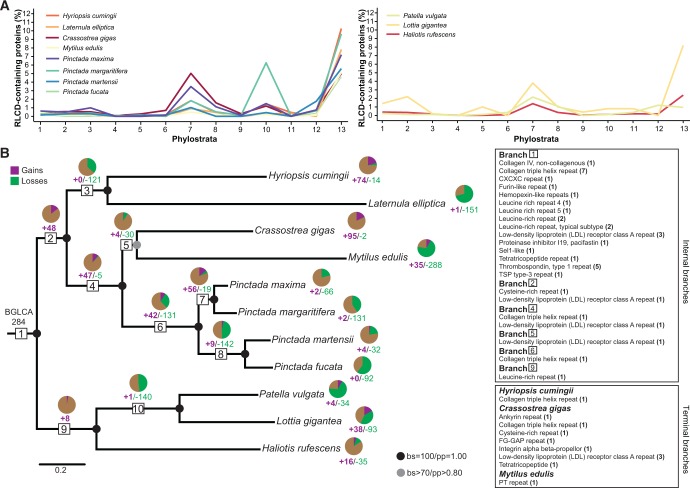
Molluscs fabricate shells of incredible diversity and complexity by localized secretions from the dorsal epithelium of the mantle. Although distantly related molluscs express remarkably different secreted gene products, it remains unclear if the evolution of shell structure and pattern is underpinned by the differential co-option of conserved genes or the integration of lineage-specific genes into the mantle regulatory program. To address this, we compare the mantle transcriptomes of 11 bivalves and gastropods of varying relatedness. We find that each species, including four Pinctada (pearl oyster) species that diverged within the last 20 Ma, expresses a unique mantle secretome. Lineage- or species-specific genes comprise a large proportion of each species' mantle secretome. A majority of these secreted proteins have unique domain architectures that include repetitive, low complexity domains (RLCDs), which evolve rapidly, and have a proclivity to expand, contract and rearrange in the genome. There are also a large number of secretome genes expressed in the mantle that arose before the origin of gastropods and bivalves. Each species expresses a unique set of these more ancient genes consistent with their independent co-option into these mantle gene regulatory networks. From this analysis, we infer lineage-specific secretomes underlie shell diversity, and include both rapidly evolving RLCD-containing proteins, and the continual recruitment and loss of both ancient and recently evolved genes into the periphery of the regulatory network controlling gene expression in the mantle epithelium.
Keywords: bivalve; co-option; gastropod; lineage-specific novelties; mantle secretome; shell formation.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
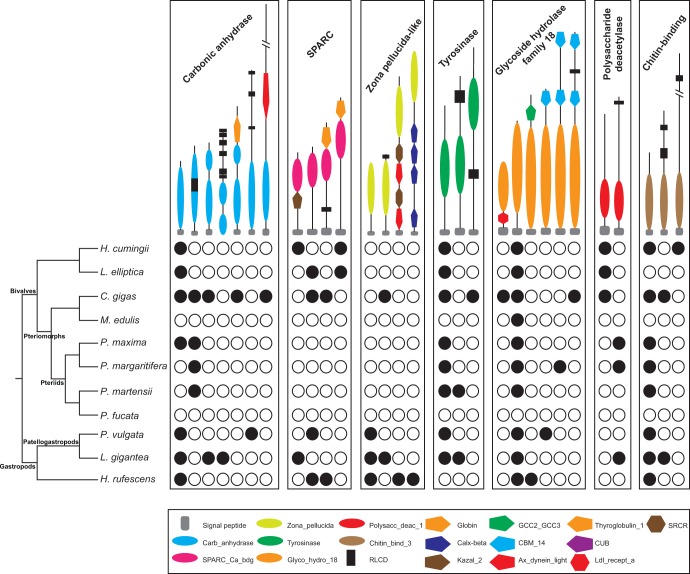
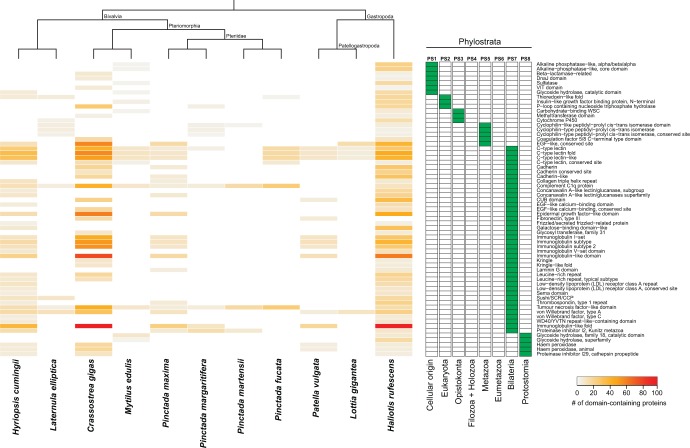
References
-
- Aguilera F, McDougall C, Degnan BM.. 2014. Evolution of the tyrosinase gene family in bivalve molluscs: independent expansion of the mantle gene repertoire. Acta Biomater. 10:3855–3865. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
