

Heat Shock Protein 90 Ensures Efficient Mumps Virus Replication by Assisting with Viral Polymerase Complex Formation
- PMID: 28053100
- PMCID: PMC5331814
- DOI: 10.1128/JVI.02220-16
Heat Shock Protein 90 Ensures Efficient Mumps Virus Replication by Assisting with Viral Polymerase Complex Formation
Abstract
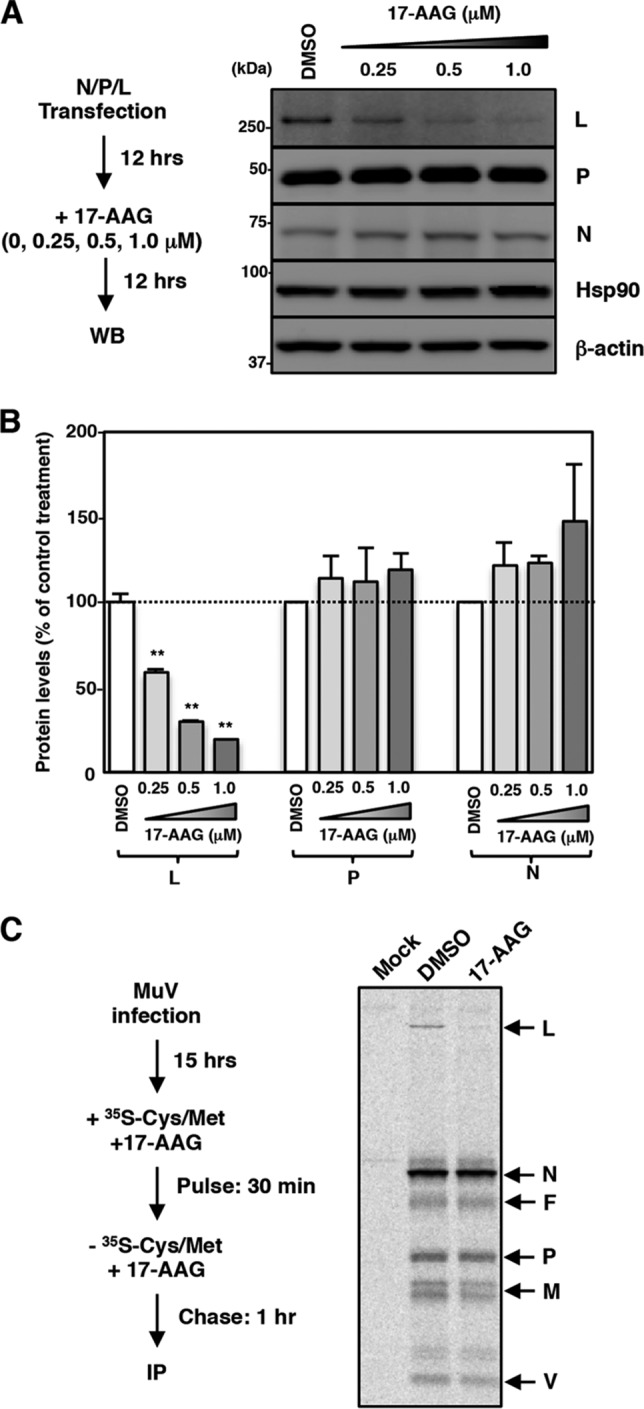
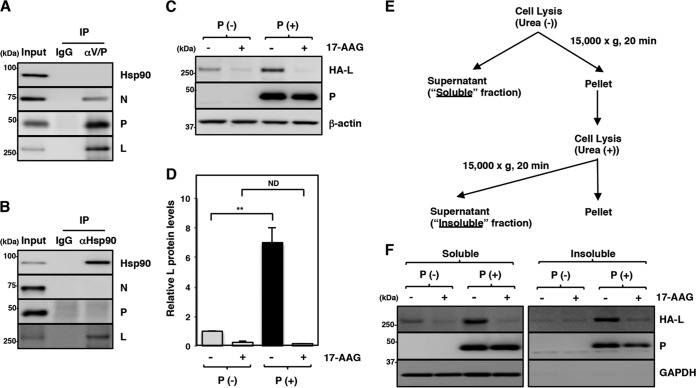
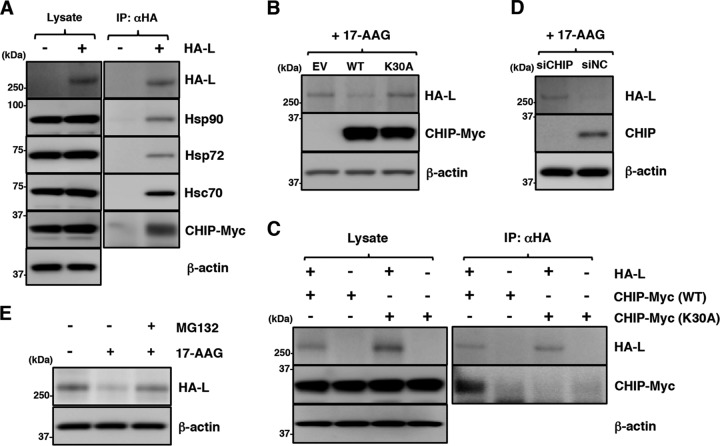
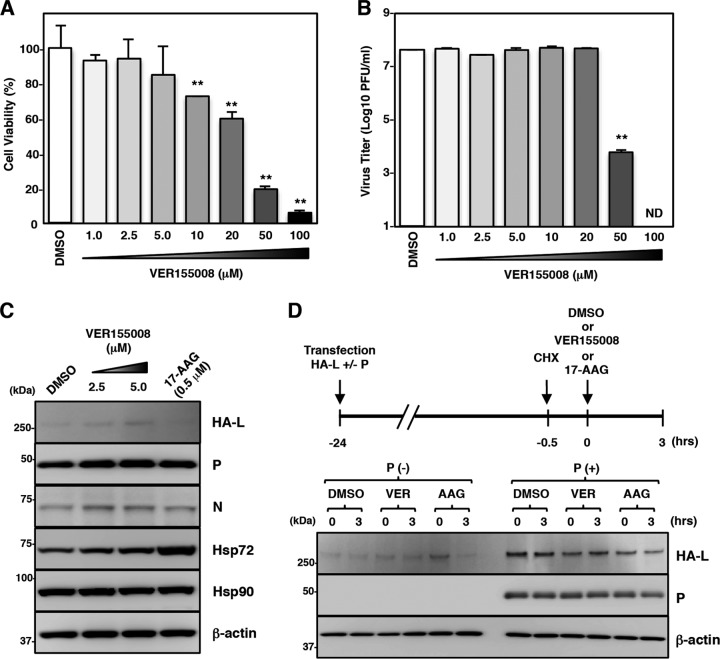
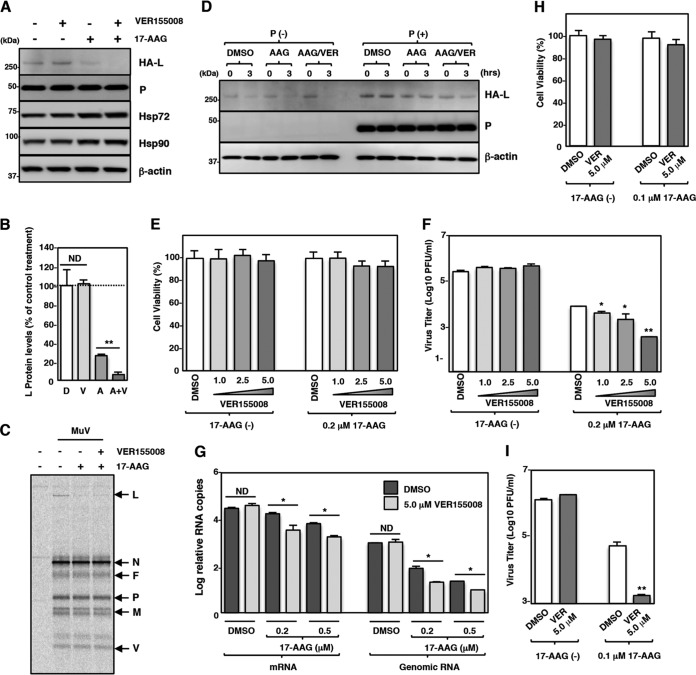
Paramyxoviral RNAs are synthesized by a viral RNA-dependent RNA polymerase (RdRp) consisting of the large (L) protein and its cofactor phosphoprotein (P protein). The L protein is a multifunctional protein that catalyzes RNA synthesis, mRNA capping, and mRNA polyadenylation. Growing evidence shows that the stability of several paramyxovirus L proteins is regulated by heat shock protein 90 (Hsp90). In this study, we demonstrated that Hsp90 activity was important for mumps virus (MuV) replication. The Hsp90 activity was required for L-protein stability and activity because an Hsp90-specific inhibitor, 17-allylamino-17-demethoxygeldanamycin (17-AAG), destabilized the MuV L protein and suppressed viral RNA synthesis. However, once the L protein formed a mature polymerase complex with the P protein, Hsp90 activity was no longer required for the stability and activity of the L protein. When the Hsp90 activity was inhibited, the MuV L protein was degraded through the CHIP (C terminus of Hsp70-interacting protein)-mediated proteasomal pathway. High concentrations of 17-AAG showed strong cytotoxicity to certain cell types, but combined use of an Hsp70 inhibitor, VER155008, potentiated degradation of the L protein, allowing a sufficient reduction of 17-AAG concentration to block MuV replication with minimum cytotoxicity. Regulation of the L protein by Hsp90 and Hsp70 chaperones was also demonstrated for another paramyxovirus, the measles virus. Collectively, our data show that the Hsp90/Hsp70 chaperone machinery assists in the maturation of the paramyxovirus L protein and thereby in the formation of a mature RdRp complex and efficient viral replication.IMPORTANCE Heat shock protein 90 (Hsp90) is nearly universally required for viral protein homeostasis. Here, we report that Hsp90 activity is required for efficient propagation of mumps virus (MuV). Hsp90 functions in the maintenance of the catalytic subunit of viral polymerase, the large (L) protein, prior to formation of a mature polymerase complex with the polymerase cofactor of L, phosphoprotein. Hsp70 collaborates with Hsp90 to regulate biogenesis of the MuV L protein. The functions of these chaperones on the viral polymerase may be common among paramyxoviruses because the L protein of measles virus is also similarly regulated. Our data provide important insights into the molecular mechanisms of paramyxovirus polymerase maturation as well as a basis for the development of novel antiviral drugs.
Keywords: heat shock protein 90; large protein; mumps virus; polymerase.
Copyright © 2017 American Society for Microbiology.
Figures



Similar articles
-
The R2TP complex regulates paramyxovirus RNA synthesis.PLoS Pathog. 2019 May 23;15(5):e1007749. doi: 10.1371/journal.ppat.1007749. eCollection 2019 May. PLoS Pathog. 2019. PMID: 31121004 Free PMC article.
-
Heat Shock Protein 90 Ensures the Integrity of Rubella Virus p150 Protein and Supports Viral Replication.J Virol. 2019 Oct 29;93(22):e01142-19. doi: 10.1128/JVI.01142-19. Print 2019 Nov 15. J Virol. 2019. PMID: 31484751 Free PMC article.
-
HSP90 Chaperoning in Addition to Phosphoprotein Required for Folding but Not for Supporting Enzymatic Activities of Measles and Nipah Virus L Polymerases.J Virol. 2016 Jul 11;90(15):6642-6656. doi: 10.1128/JVI.00602-16. Print 2016 Aug 1. J Virol. 2016. PMID: 27170753 Free PMC article.
-
Role of Heat Shock Proteins (HSP70 and HSP90) in Viral Infection.Int J Mol Sci. 2021 Aug 29;22(17):9366. doi: 10.3390/ijms22179366. Int J Mol Sci. 2021. PMID: 34502274 Free PMC article. Review.
-
Chaperoning the Mononegavirales: Current Knowledge and Future Directions.Viruses. 2018 Dec 8;10(12):699. doi: 10.3390/v10120699. Viruses. 2018. PMID: 30544818 Free PMC article. Review.
Cited by
-
Proteostasis in Viral Infection: Unfolding the Complex Virus-Chaperone Interplay.Cold Spring Harb Perspect Biol. 2020 Mar 2;12(3):a034090. doi: 10.1101/cshperspect.a034090. Cold Spring Harb Perspect Biol. 2020. PMID: 30858229 Free PMC article. Review.
-
Molecular mechanisms of stress-induced reactivation in mumps virus condensates.Cell. 2023 Apr 27;186(9):1877-1894.e27. doi: 10.1016/j.cell.2023.03.015. Cell. 2023. PMID: 37116470 Free PMC article.
-
A Novel Role for the Regulatory Nod-Like Receptor NLRP12 in Anti-Dengue Virus Response.Front Immunol. 2021 Dec 9;12:744880. doi: 10.3389/fimmu.2021.744880. eCollection 2021. Front Immunol. 2021. PMID: 34956178 Free PMC article.
-
How order and disorder within paramyxoviral nucleoproteins and phosphoproteins orchestrate the molecular interplay of transcription and replication.Cell Mol Life Sci. 2017 Sep;74(17):3091-3118. doi: 10.1007/s00018-017-2556-3. Epub 2017 Jun 9. Cell Mol Life Sci. 2017. PMID: 28600653 Free PMC article. Review.
-
SNARE protein USE1 is involved in the glycosylation and the expression of mumps virus fusion protein and important for viral propagation.PLoS Pathog. 2022 Dec 8;18(12):e1010949. doi: 10.1371/journal.ppat.1010949. eCollection 2022 Dec. PLoS Pathog. 2022. PMID: 36480520 Free PMC article.
References
-
- Lamb RA, Parks GD. 2006. Paramyxoviridae: the viruses and their replication, p 1449–1496. In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Whelan SP, Barr JN, Wertz GW. 2004. Transcription and replication of nonsegmented negative-strand RNA viruses. Curr Top Microbiol Immunol 283:61–119. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
