

ADAR2 regulates RNA stability by modifying access of decay-promoting RNA-binding proteins
- PMID: 28053121
- PMCID: PMC5397167
- DOI: 10.1093/nar/gkw1304
ADAR2 regulates RNA stability by modifying access of decay-promoting RNA-binding proteins
Abstract
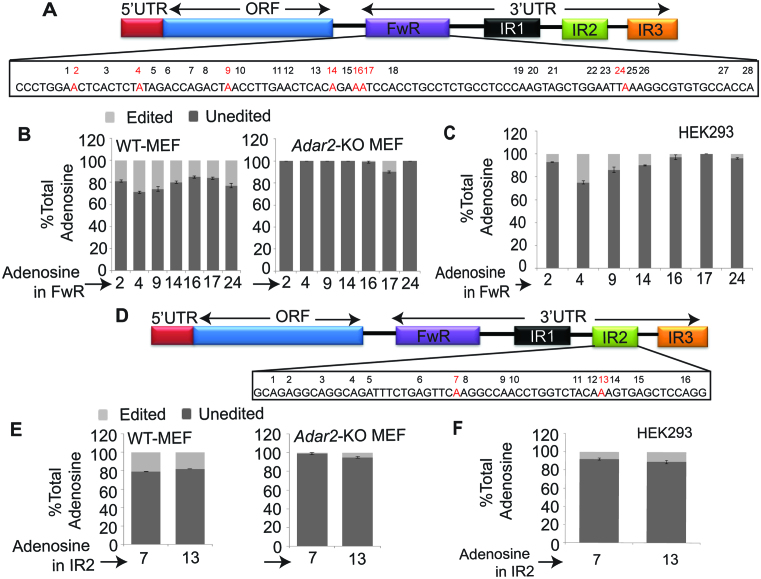
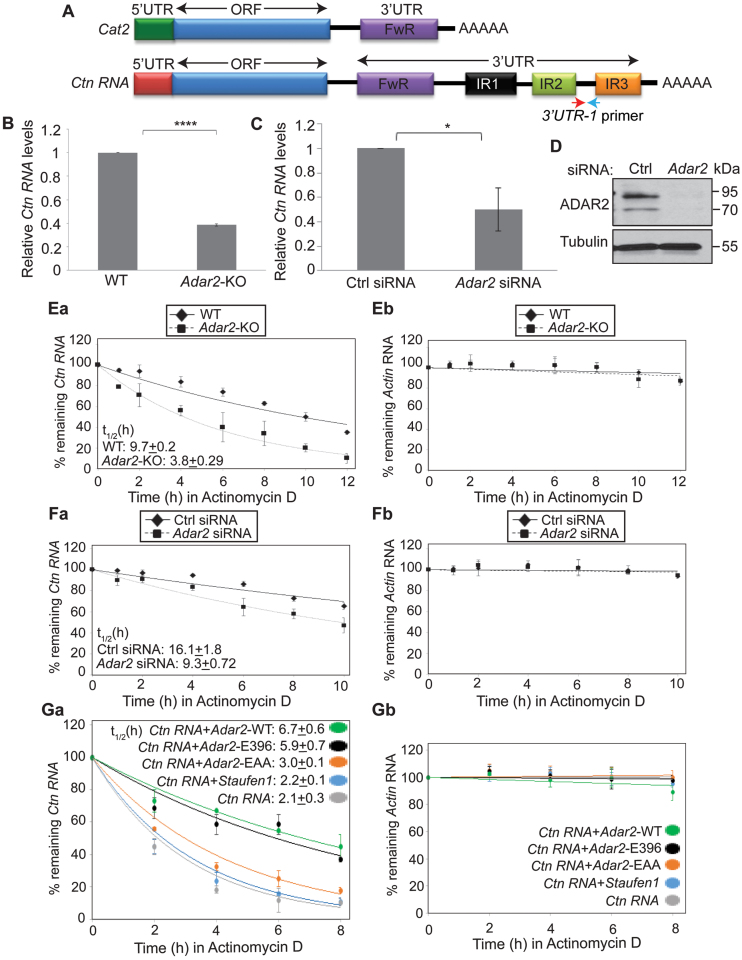
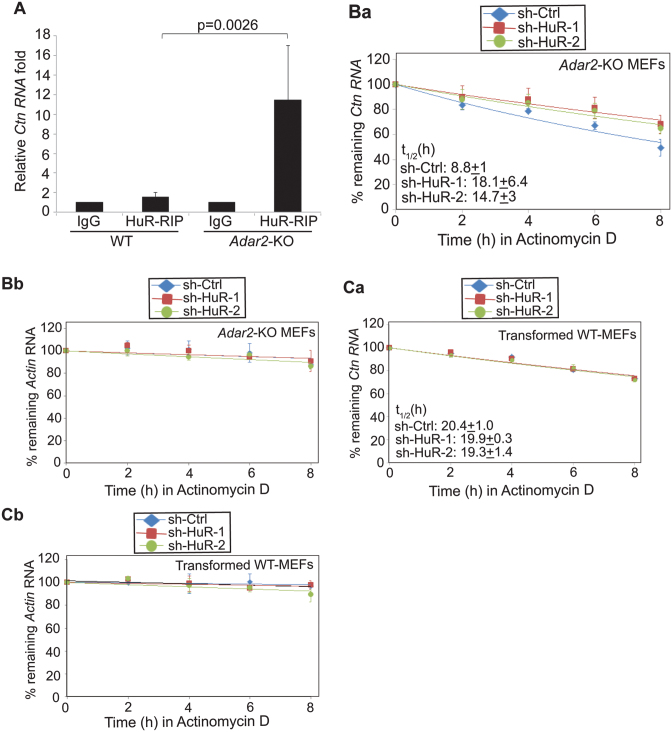
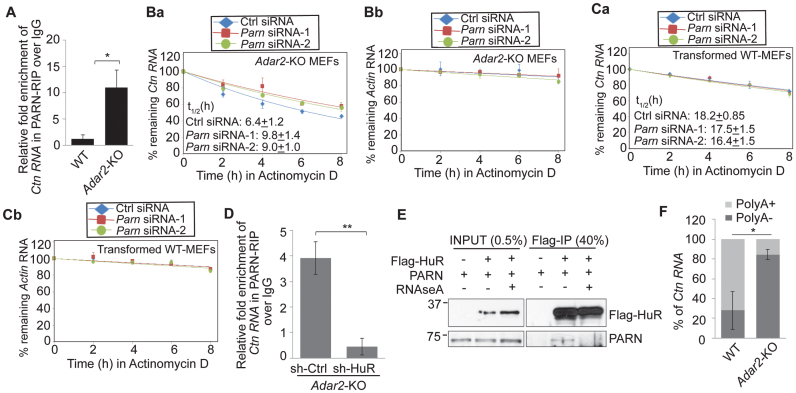
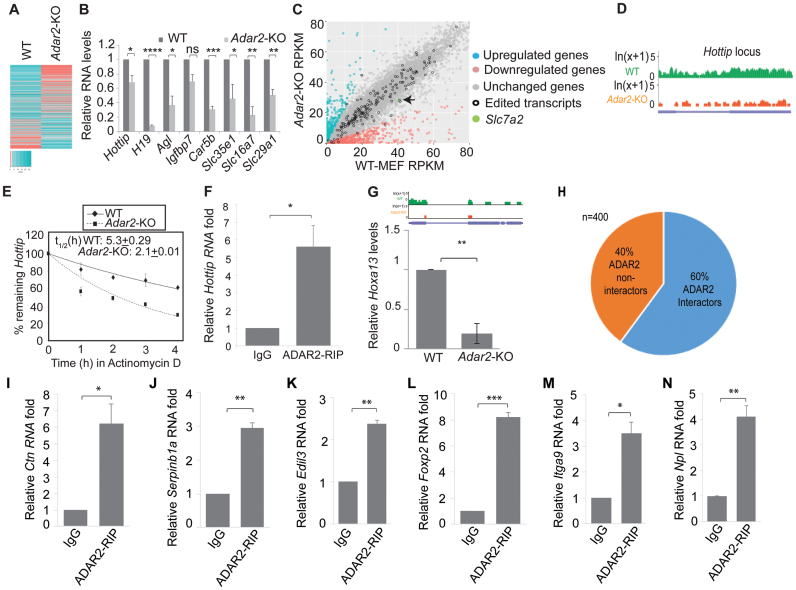
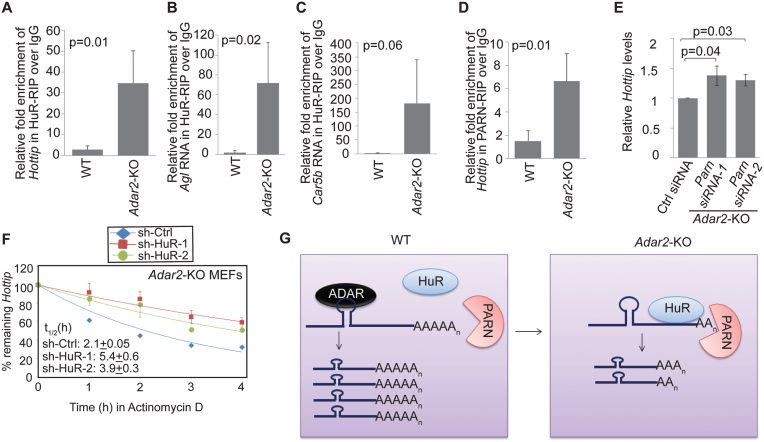
Adenosine deaminases acting on RNA (ADARs) catalyze the editing of adenosine residues to inosine (A-to-I) within RNA sequences, mostly in the introns and UTRs (un-translated regions). The significance of editing within non-coding regions of RNA is poorly understood. Here, we demonstrate that association of ADAR2 with RNA stabilizes a subset of transcripts. ADAR2 interacts with and edits the 3΄UTR of nuclear-retained Cat2 transcribed nuclear RNA (Ctn RNA). In absence of ADAR2, the abundance and half-life of Ctn RNA are significantly reduced. Furthermore, ADAR2-mediated stabilization of Ctn RNA occurred in an editing-independent manner. Unedited Ctn RNA shows enhanced interaction with the RNA-binding proteins HuR and PARN [Poly(A) specific ribonuclease deadenylase]. HuR and PARN destabilize Ctn RNA in absence of ADAR2, indicating that ADAR2 stabilizes Ctn RNA by antagonizing its degradation by PARN and HuR. Transcriptomic analysis identified other RNAs that are regulated by a similar mechanism. In summary, we identify a regulatory mechanism whereby ADAR2 enhances target RNA stability by limiting the interaction of RNA-destabilizing proteins with their cognate substrates.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Hartner J.C., Schmittwolf C., Kispert A., Muller A.M., Higuchi M., Seeburg P.H.. Liver disintegration in the mouse embryo caused by deficiency in the RNA-editing enzyme ADAR1. J. Biol. Chem. 2004; 279:4894–4902. - PubMed
-
- Higuchi M., Maas S., Single F.N., Hartner J., Rozov A., Burnashev N., Feldmeyer D., Sprengel R., Seeburg P.H.. Point mutation in an AMPA receptor gene rescues lethality in mice deficient in the RNA-editing enzyme ADAR2. Nature. 2000; 406:78–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
