

Macroautophagy and Selective Mitophagy Ameliorate Chondrogenic Differentiation Potential in Adipose Stem Cells of Equine Metabolic Syndrome: New Findings in the Field of Progenitor Cells Differentiation
- PMID: 28053691
- PMCID: PMC5178365
- DOI: 10.1155/2016/3718468
Macroautophagy and Selective Mitophagy Ameliorate Chondrogenic Differentiation Potential in Adipose Stem Cells of Equine Metabolic Syndrome: New Findings in the Field of Progenitor Cells Differentiation
Erratum in
-
Corrigendum to "Macroautophagy and Selective Mitophagy Ameliorate Chondrogenic Differentiation Potential in Adipose Stem Cells of Equine Metabolic Syndrome: New Findings in the Field of Progenitor Cells Differentiation".Oxid Med Cell Longev. 2017;2017:3861790. doi: 10.1155/2017/3861790. Epub 2017 Jul 27. Oxid Med Cell Longev. 2017. PMID: 28831295 Free PMC article.
Abstract
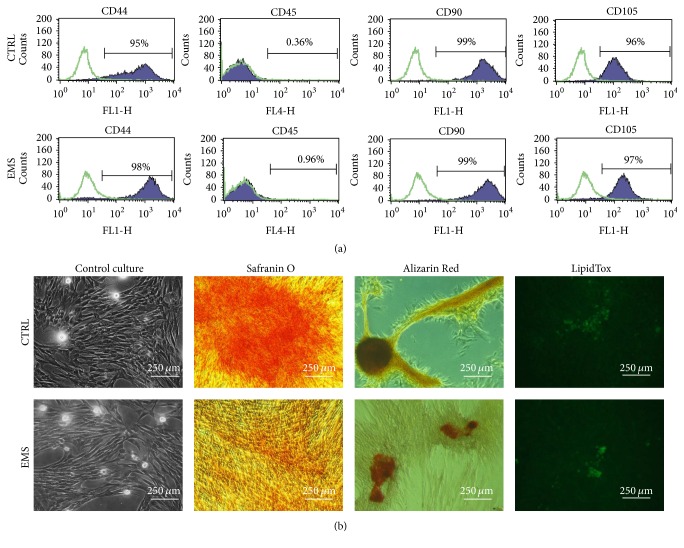
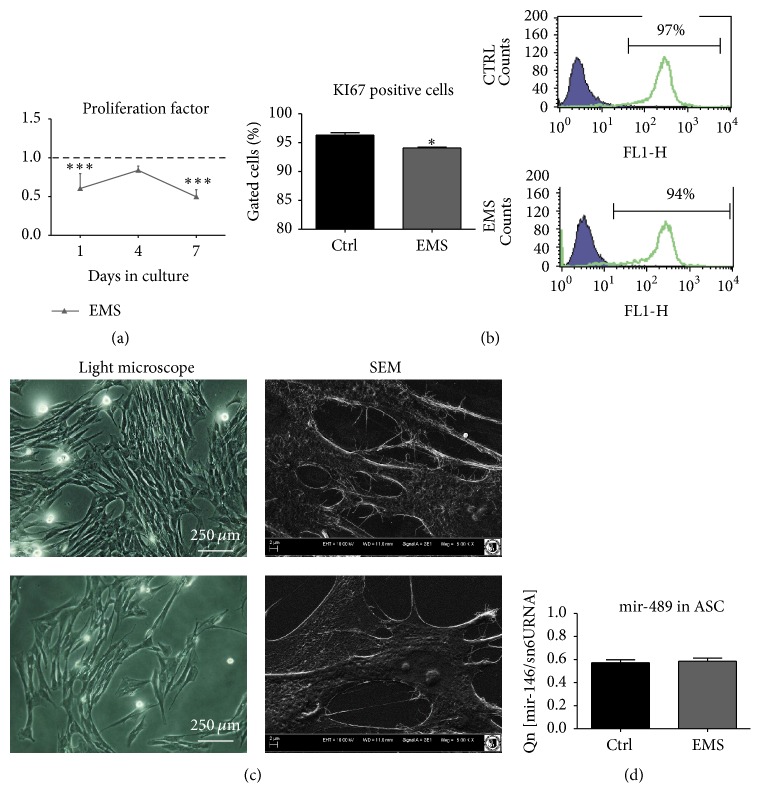
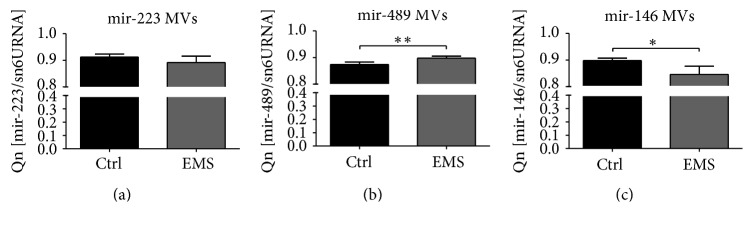
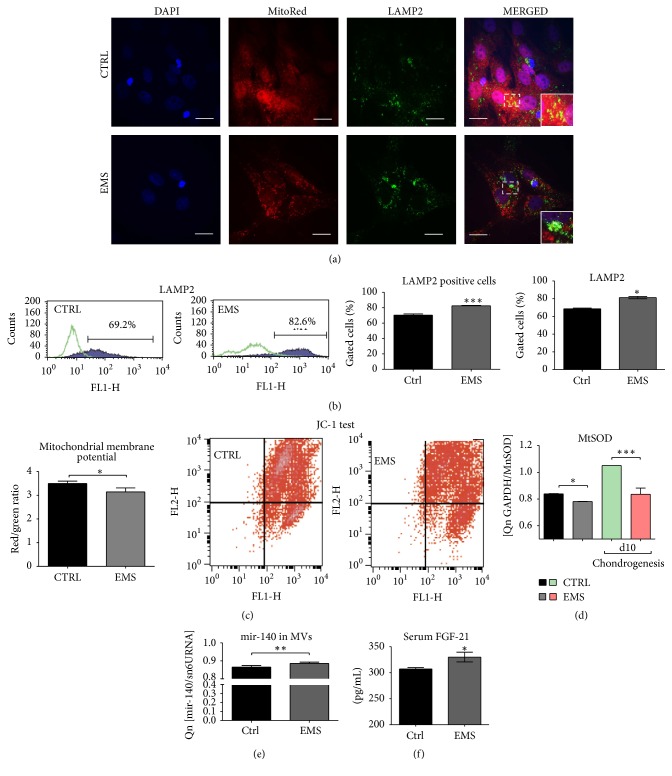
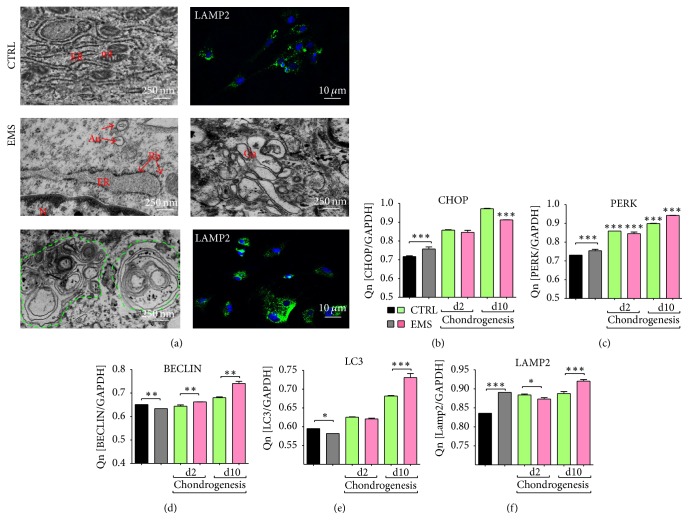
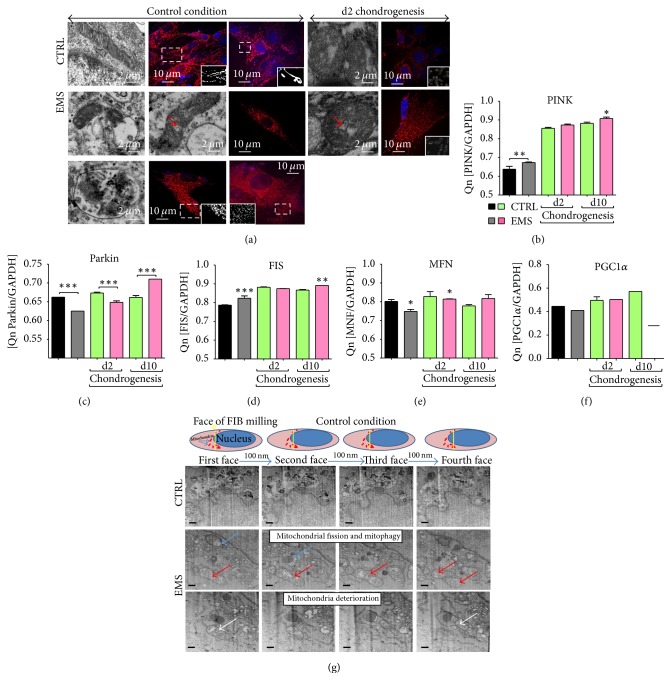
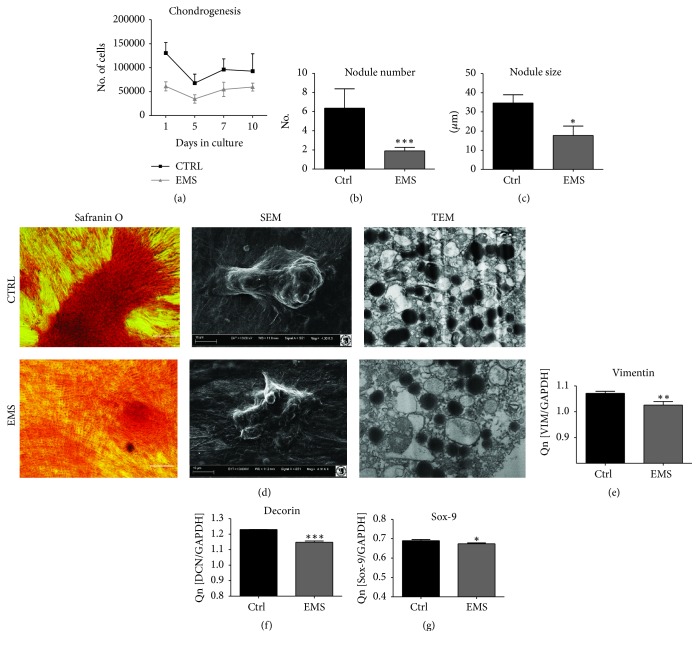
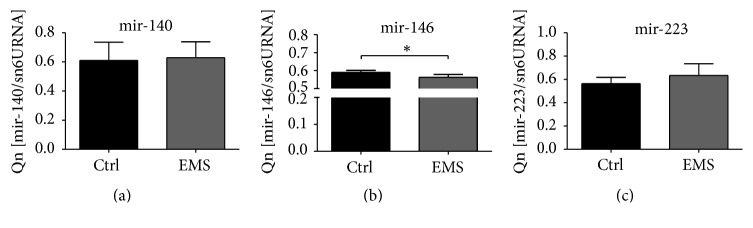
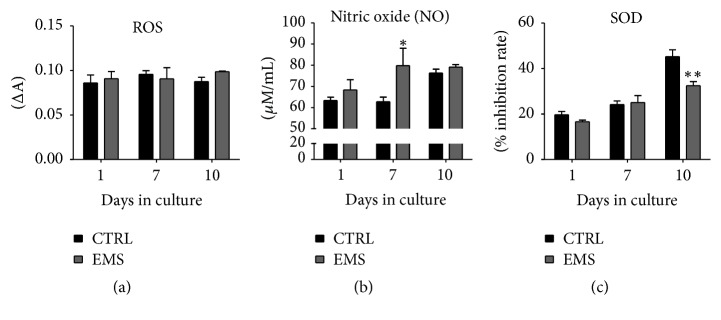
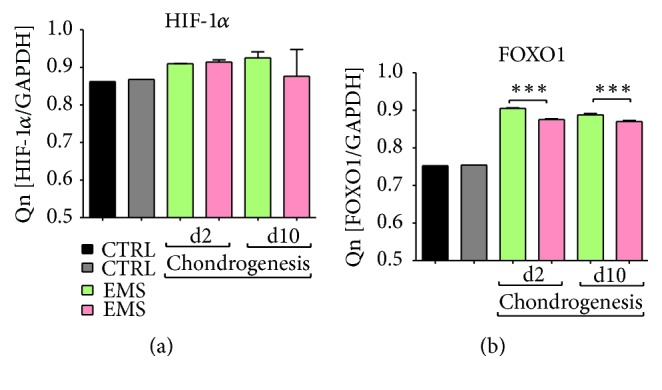
Equine metabolic syndrome (EMS) is mainly characterized by insulin resistance, obesity, and local or systemic inflammation. That unfriendly environment of adipose tissue has huge impact on stem cells population (ASC) residing within. In the present study, using molecular biology techniques and multiple imaging techniques (SEM, FIB-SEM, and confocal microscopy), we evaluated the impact of EMS on ASC viability and chondrogenic differentiation. Moreover, we visualized the mitochondrial network and dynamics in ASCCTRL and ASCEMS during control and chondrogenic conditions. In control conditions, ASCEMS were characterized by increased mitochondrial fission in comparison to ASCCTRL. We found that extensive remodeling of mitochondrial network including fusion and fission occurs during early step of differentiation. Moreover, we observed mitochondria morphology deterioration in ASCEMS. These conditions seem to cause autophagic shift in ASCEMS, as we observed increased accumulation of LAMP2 and formation of multiple autophagosomes in those cells, some of which contained dysfunctional mitochondria. "Autophagic" switch may be a rescue mechanism allowing ASCEMS to clear impaired by ROS proteins and mitochondria. Moreover it provides a precursors-to-macromolecules synthesis, especially during chondrogenesis. Our data indicates that autophagy in ASCEMS would be crucial for the quality control mechanisms and maintenance of cellular homeostasis ASCEMS allowing them to be in "stemness" status.
Conflict of interest statement
The authors declare that there is no conflict of interests.
Figures
References
-
- Frank N. Equine metabolic syndrome. Journal of Equine Veterinary Science. 2009;29(5):259–267. doi: 10.1016/j.jevs.2009.04.183. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
