

Genetic conflicts: the usual suspects and beyond
- PMID: 28057823
- PMCID: PMC5278622
- DOI: 10.1242/jeb.148148
Genetic conflicts: the usual suspects and beyond
Abstract
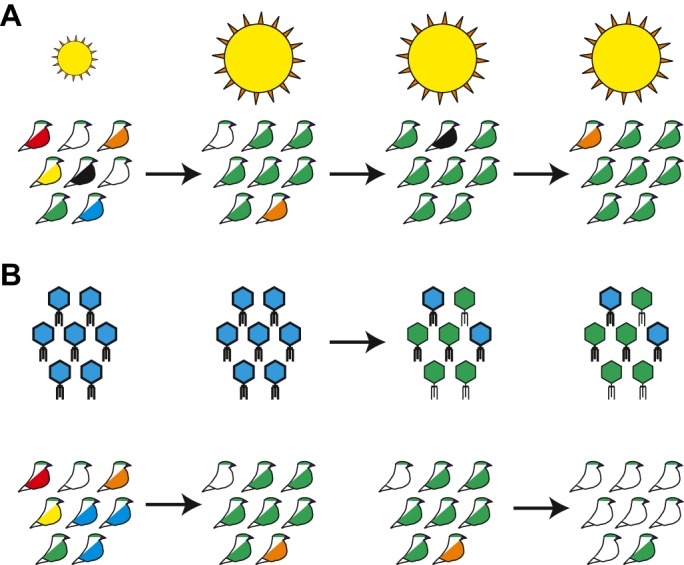
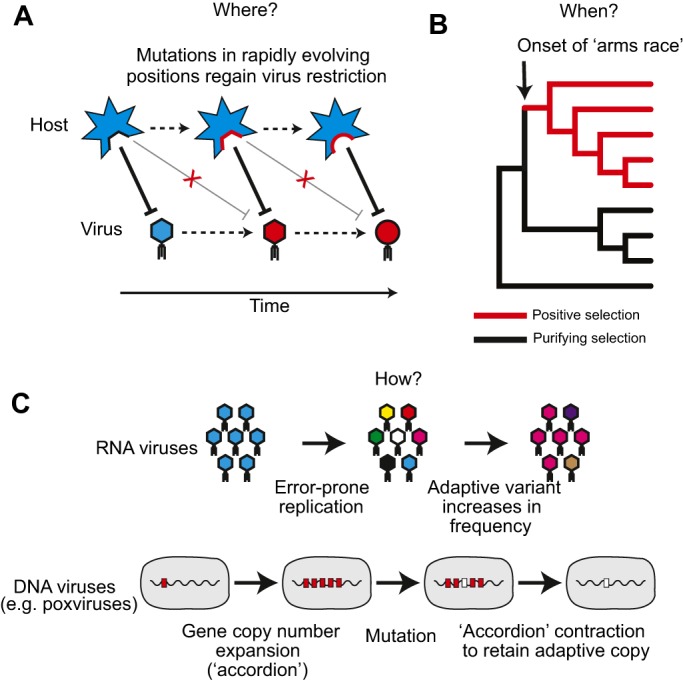
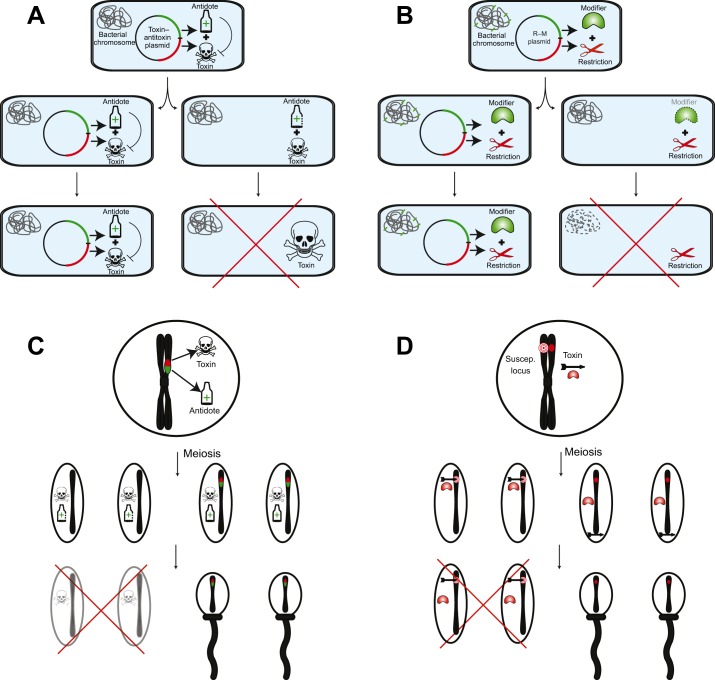
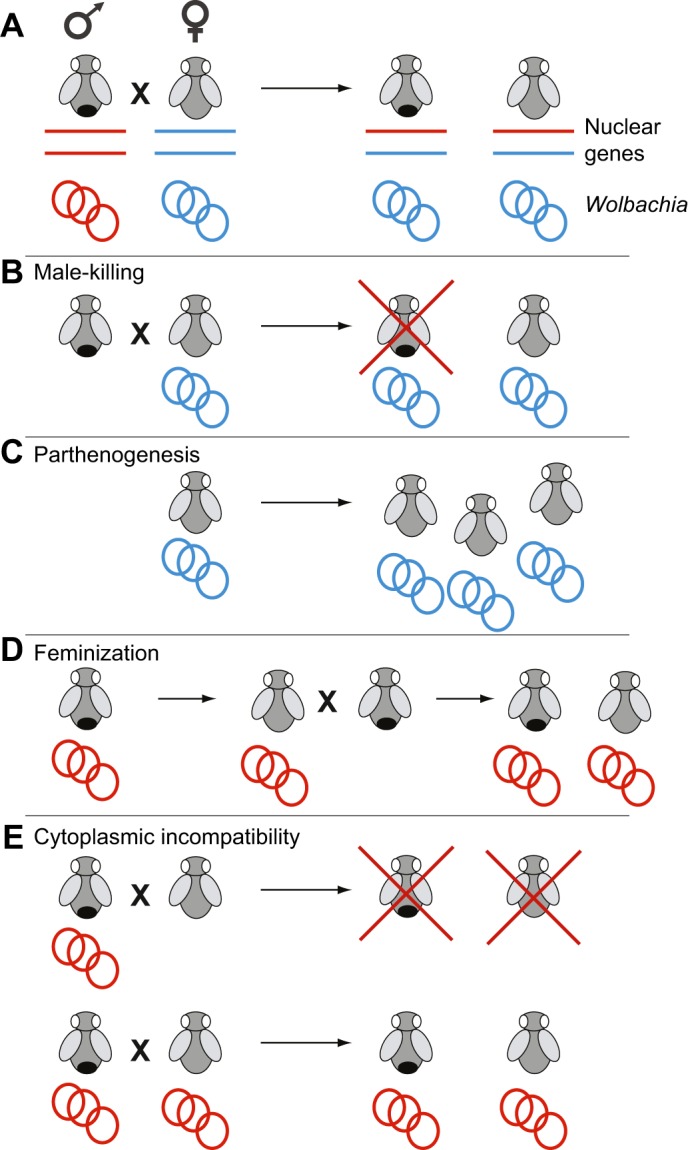
Selfishness is pervasive and manifests at all scales of biology, from societies, to individuals, to genetic elements within a genome. The relentless struggle to seek evolutionary advantages drives perpetual cycles of adaptation and counter-adaptation, commonly referred to as Red Queen interactions. In this review, we explore insights gleaned from molecular and genetic studies of such genetic conflicts, both extrinsic (between genomes) and intrinsic (within genomes or cells). We argue that many different characteristics of selfish genetic elements can be distilled into two types of advantages: an over-replication advantage (e.g. mobile genetic elements in genomes) and a transmission distortion advantage (e.g. meiotic drivers in populations). These two general categories may help classify disparate types of selfish genetic elements.
Keywords: Antagonism; Host–pathogen; Meiosis; Mitochondria; Mutualism; Red Queen; Toxin–antitoxin; Wolbachia.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
