

Deletional tolerance prevents AQP4-directed autoimmunity in mice
- PMID: 28058717
- PMCID: PMC5359142
- DOI: 10.1002/eji.201646855
Deletional tolerance prevents AQP4-directed autoimmunity in mice
Abstract
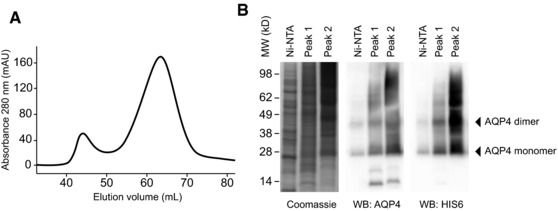
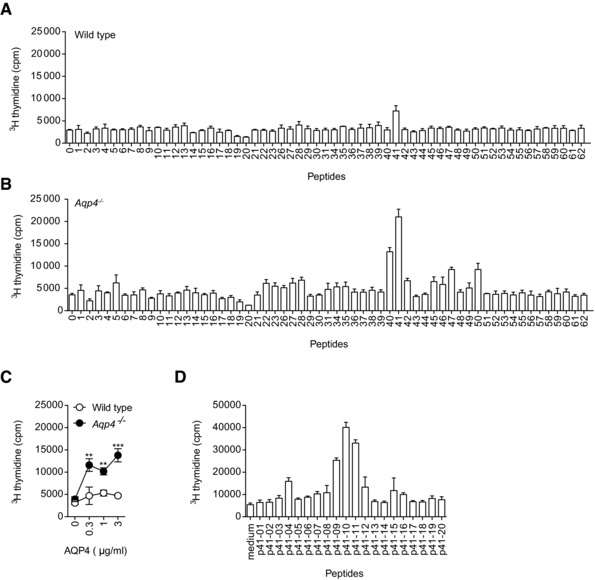
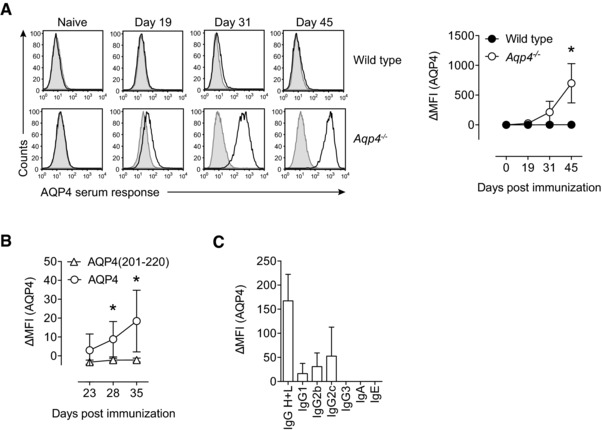
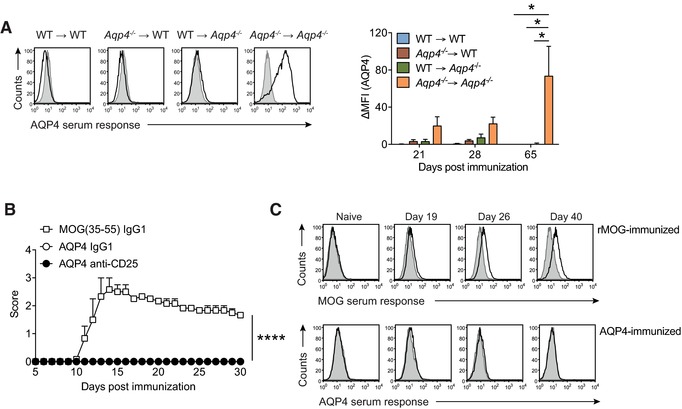
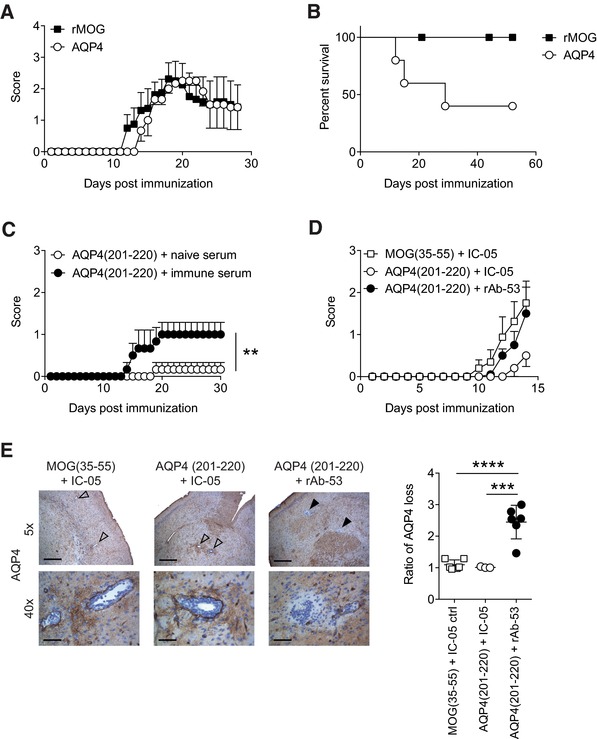
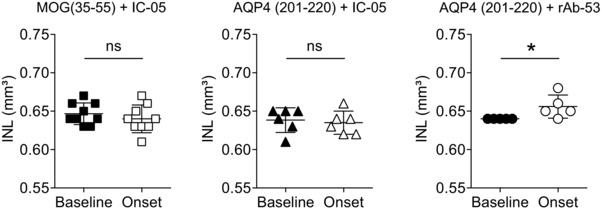
Neuromyelitis optica (NMO) is an autoimmune disorder of the central nervous system (CNS) mediated by antibodies to the water channel protein AQP4 expressed in astrocytes. The contribution of AQP4-specific T cells to the class switch recombination of pathogenic AQP4-specific antibodies and the inflammation of the blood-brain barrier is incompletely understood, as immunogenic naturally processed T-cell epitopes of AQP4 are unknown. By immunizing Aqp4-/- mice with full-length murine AQP4 protein followed by recall with overlapping peptides, we here identify AQP4(201-220) as the major immunogenic IAb -restricted epitope of AQP4. We show that WT mice do not harbor AQP4(201-220)-specific T-cell clones in their natural repertoire due to deletional tolerance. However, immunization with AQP4(201-220) of Rag1-/- mice reconstituted with the mature T-cell repertoire of Aqp4-/- mice elicits an encephalomyelitic syndrome. Similarly to the T-cell repertoire, the B-cell repertoire of WT mice is "purged" of AQP4-specific B cells, and robust serum responses to AQP4 are only mounted in Aqp4-/- mice. While AQP4(201-220)-specific T cells alone induce encephalomyelitis, NMO-specific lesional patterns in the CNS and the retina only occur in the additional presence of anti-AQP4 antibodies. Thus, failure of deletional T-cell and B-cell tolerance against AQP4 is a prerequisite for clinically manifest NMO.
Keywords: Aquaporin 4 (AQP4); B cell; Encephalitogenic epitope; Experimental neuromyelitis optica; Optical coherence tomography; T cell; Tolerance.
© 2017 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
References
-
- Lennon, V. A. , Wingerchuk, D. M. , Kryzer, T. J. , Pittock, S. J. , Lucchinetti, C. F. , Fujihara, K. , Nakashima, I. et al., A serum autoantibody marker of neuromyelitis optica: distinction from multiple sclerosis. Lancet 2004. 364: 2106–2112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
