

Control of meiotic pairing and recombination by chromosomally tethered 26S proteasome
- PMID: 28059715
- PMCID: PMC6054871
- DOI: 10.1126/science.aaf4778
Control of meiotic pairing and recombination by chromosomally tethered 26S proteasome
Abstract
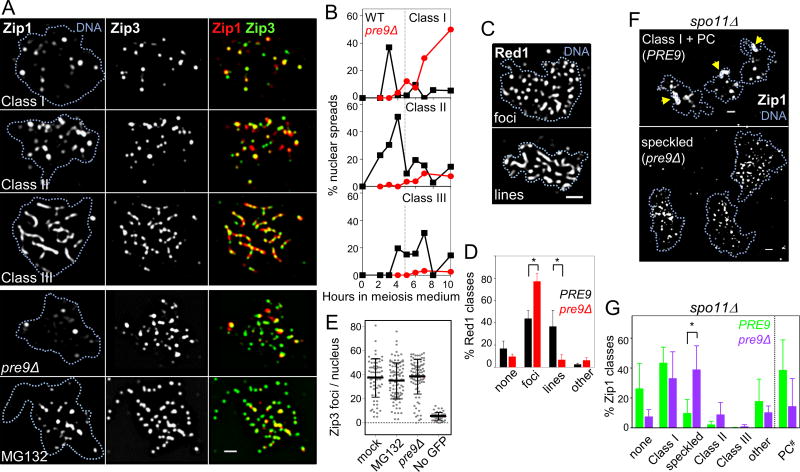
During meiosis, paired homologous chromosomes (homologs) become linked via the synaptonemal complex (SC) and crossovers. Crossovers mediate homolog segregation and arise from self-inflicted double-strand breaks (DSBs). Here, we identified a role for the proteasome, the multisubunit protease that degrades proteins in the nucleus and cytoplasm, in homolog juxtaposition and crossing over. Without proteasome function, homologs failed to pair and instead remained associated with nonhomologous chromosomes. Although dispensable for noncrossover formation, a functional proteasome was required for a coordinated transition that entails SC assembly between longitudinally organized chromosome axes and stable strand exchange of crossover-designated DSBs. Notably, proteolytic core and regulatory proteasome particles were recruited to chromosomes by Zip3, the ortholog of mammalian E3 ligase RNF212, and SC protein Zip1 . We conclude that proteasome functions along meiotic chromosomes are evolutionarily conserved.
Copyright © 2017, American Association for the Advancement of Science.
Figures



Comment in
-
When degradation spurs segregation.Science. 2017 Jan 27;355(6323):349-350. doi: 10.1126/science.aam5904. Science. 2017. PMID: 28126771 No abstract available.
-
Proteasomes on the chromosome.Cell Res. 2017 May;27(5):602-603. doi: 10.1038/cr.2017.28. Epub 2017 Mar 7. Cell Res. 2017. PMID: 28266542 Free PMC article.
References
-
- Tsubouchi T, Roeder GS. A synaptonemal complex protein promotes homology-independent centromere coupling. Science. 2005;308:870–873. - PubMed
-
- Börner GV, Kleckner N, Hunter N. Crossover/ noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell. 2004;117:29–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
