

KCTD2, an adaptor of Cullin3 E3 ubiquitin ligase, suppresses gliomagenesis by destabilizing c-Myc
- PMID: 28060381
- PMCID: PMC5384019
- DOI: 10.1038/cdd.2016.151
KCTD2, an adaptor of Cullin3 E3 ubiquitin ligase, suppresses gliomagenesis by destabilizing c-Myc
Abstract
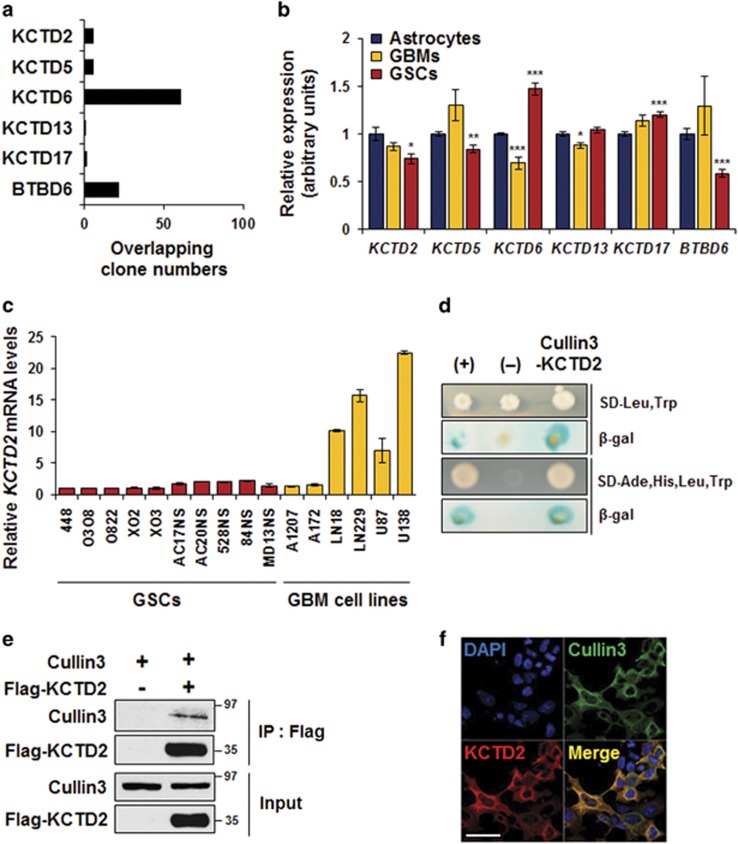
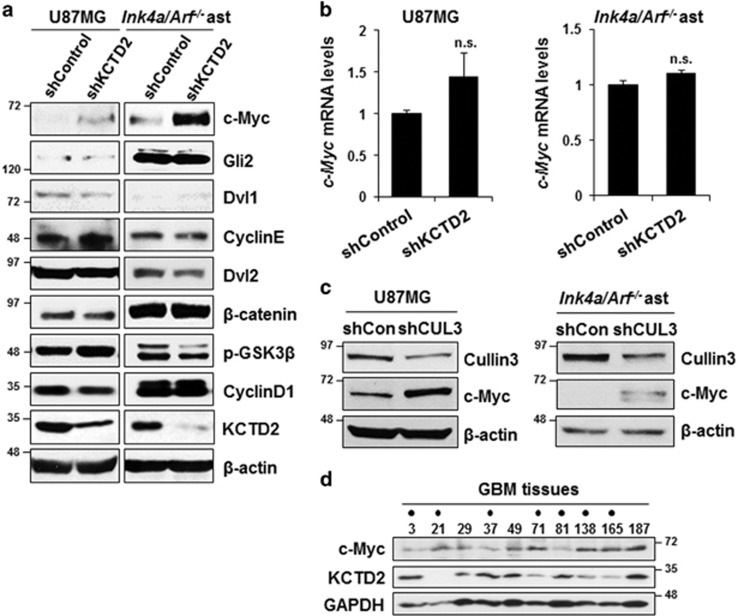
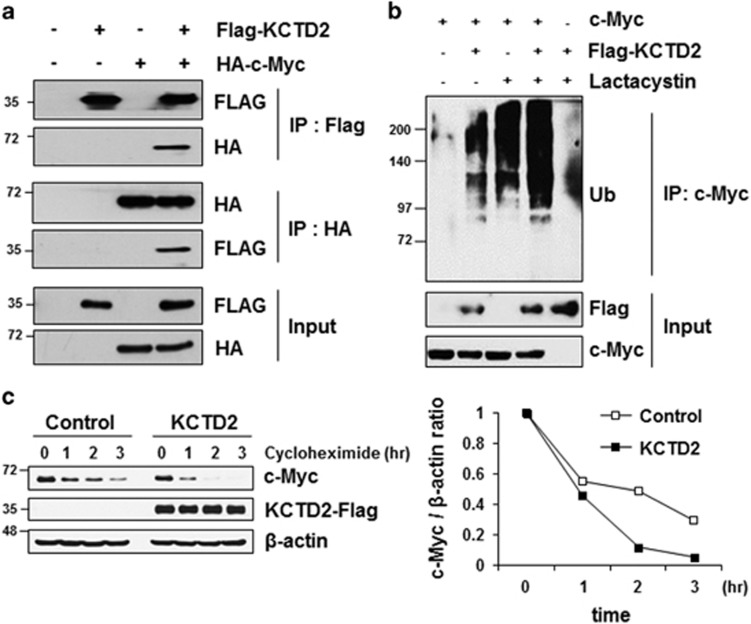
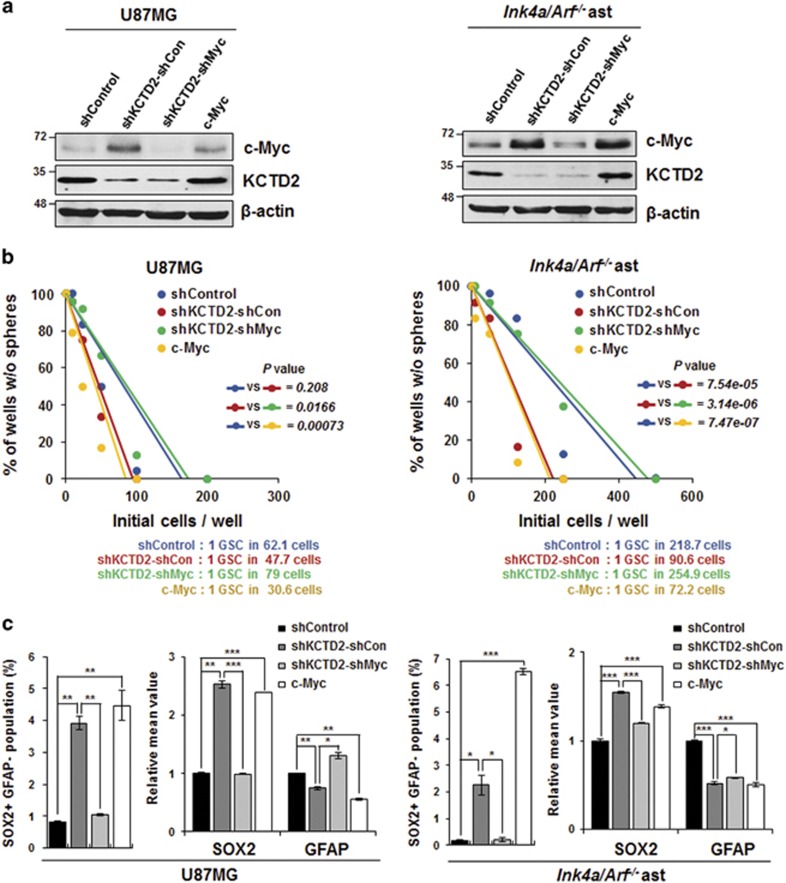
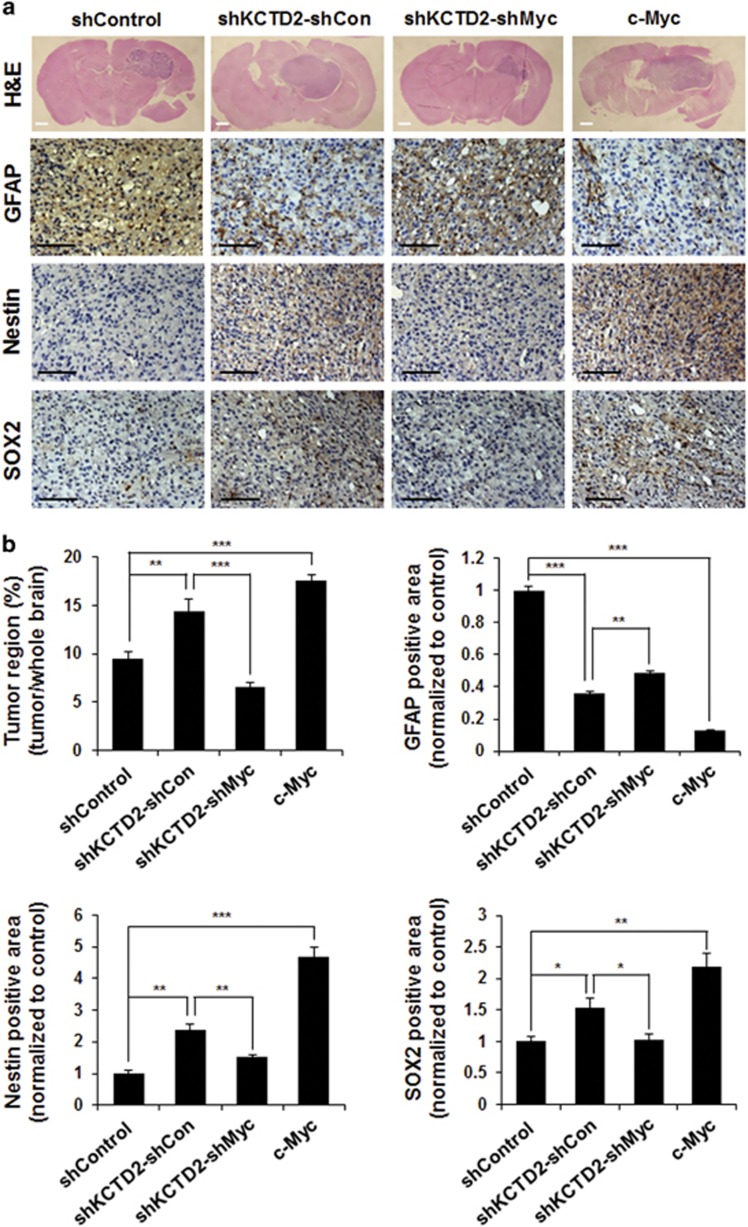
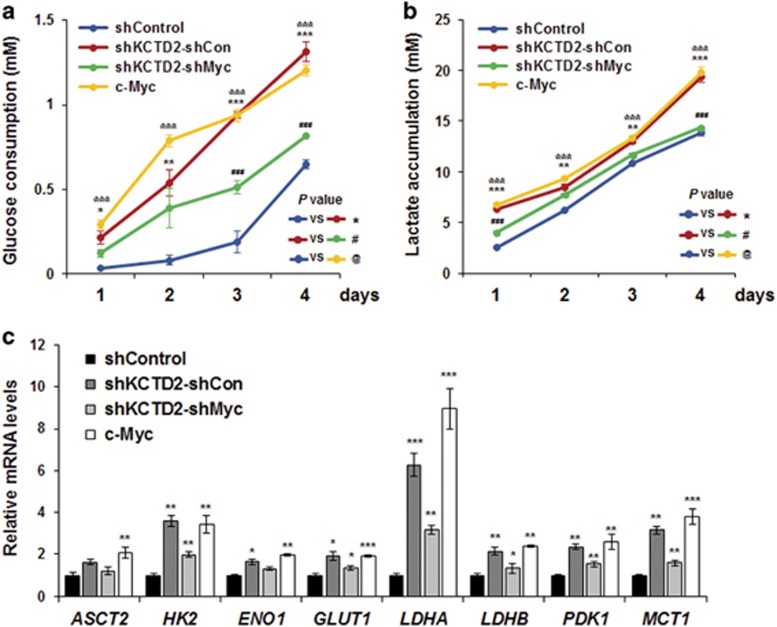
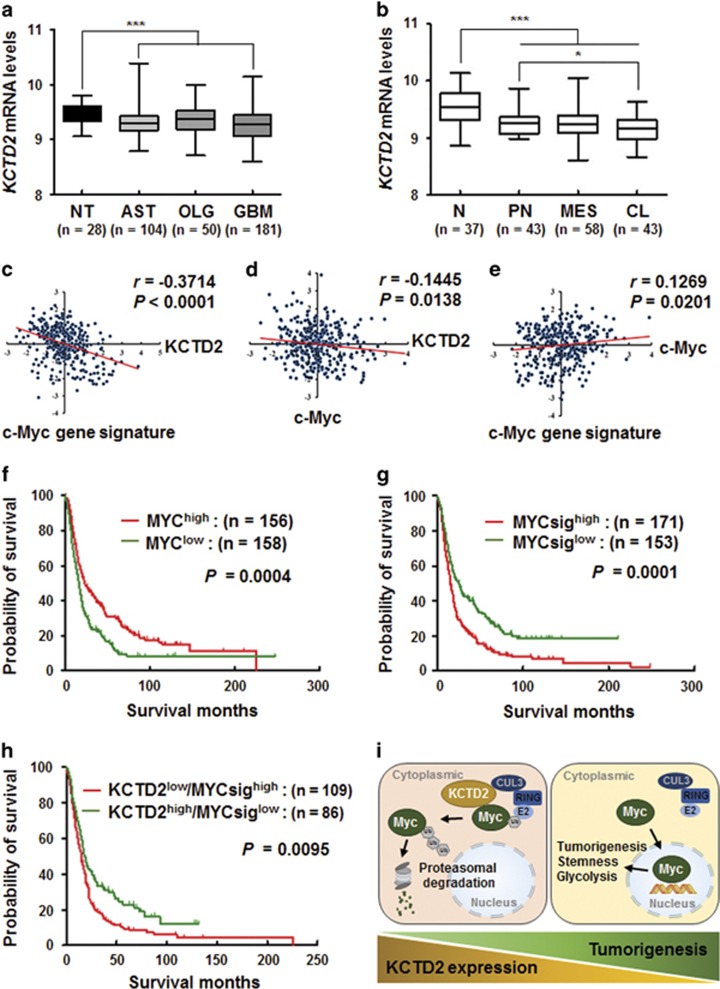
Cullin3 E3 ubiquitin ligase ubiquitinates a wide range of substrates through substrate-specific adaptors Bric-a-brac, Tramtrack, and Broad complex (BTB) domain proteins. These E3 ubiquitin ligase complexes are involved in diverse cellular functions. Our recent study demonstrated that decreased Cullin3 expression induces glioma initiation and correlates with poor prognosis of patients with malignant glioma. However, the substrate recognition mechanism associated with tumorigenesis is not completely understood. Through yeast two-hybrid screening, we identified potassium channel tetramerization domain-containing 2 (KCTD2) as a BTB domain protein that binds to Cullin3. The interaction of Cullin3 and KCTD2 was verified using immunoprecipitation and immunofluorescence. Of interest, KCTD2 expression was markedly decreased in patient-derived glioma stem cells (GSCs) compared with non-stem glioma cells. Depletion of KCTD2 using a KCTD2-specific short-hairpin RNA in U87MG glioma cells and primary Ink4a/Arf-deficient murine astrocytes markedly increased self-renewal activity in addition with an increased expression of stem cell markers, and mouse in vivo intracranial tumor growth. As an underlying mechanism for these KCTD2-mediated phenotypic changes, we demonstrated that KCTD2 interacts with c-Myc, which is a key stem cell factor, and causes c-Myc protein degradation by ubiquitination. As a result, KCTD2 depletion acquires GSC features and affects aerobic glycolysis via expression changes in glycolysis-associated genes through c-Myc protein regulation. Of clinical significance was our finding that patients having a profile of KCTD2 mRNA-low and c-Myc gene signature-high, but not KCTD2 mRNA-low and c-Myc mRNA-high, are strongly associated with poor prognosis. This study describes a novel regulatory mode of c-Myc protein in malignant gliomas and provides a potential framework for glioma therapy by targeting c-Myc function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
