

Watching Proteins Wiggle: Mapping Structures with Two-Dimensional Infrared Spectroscopy
- PMID: 28060489
- PMCID: PMC5500453
- DOI: 10.1021/acs.chemrev.6b00582
Watching Proteins Wiggle: Mapping Structures with Two-Dimensional Infrared Spectroscopy
Abstract
Proteins exhibit structural fluctuations over decades of time scales. From the picosecond side chain motions to aggregates that form over the course of minutes, characterizing protein structure over these vast lengths of time is important to understanding their function. In the past 15 years, two-dimensional infrared spectroscopy (2D IR) has been established as a versatile tool that can uniquely probe proteins structures on many time scales. In this review, we present some of the basic principles behind 2D IR and show how they have, and can, impact the field of protein biophysics. We highlight experiments in which 2D IR spectroscopy has provided structural and dynamical data that would be difficult to obtain with more standard structural biology techniques. We also highlight technological developments in 2D IR that continue to expand the scope of scientific problems that can be accessed in the biomedical sciences.
Conflict of interest statement
The authors declare the following competing financial interest(s): M.T.Z. is a co-owner of PhaseTech Spectroscopy, which manufactures mid-IR pulse shapers and 2D IR spectrometers.
Figures
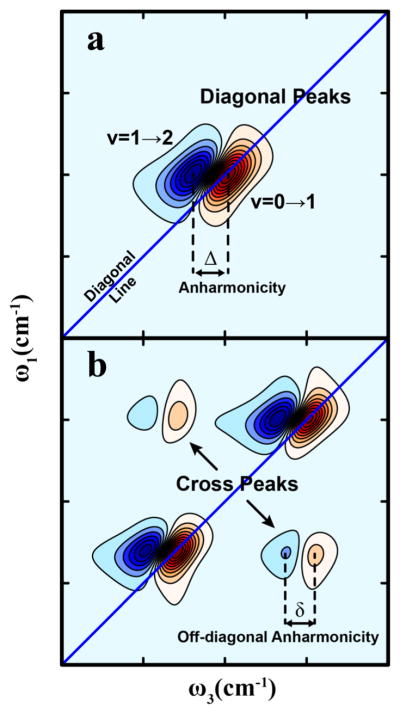
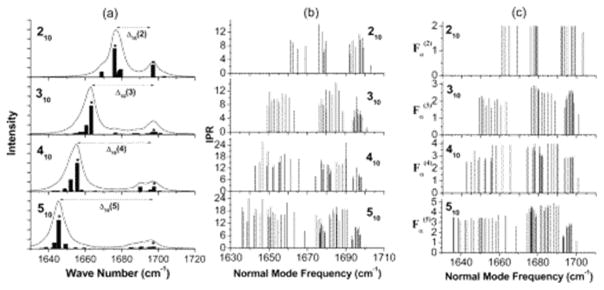
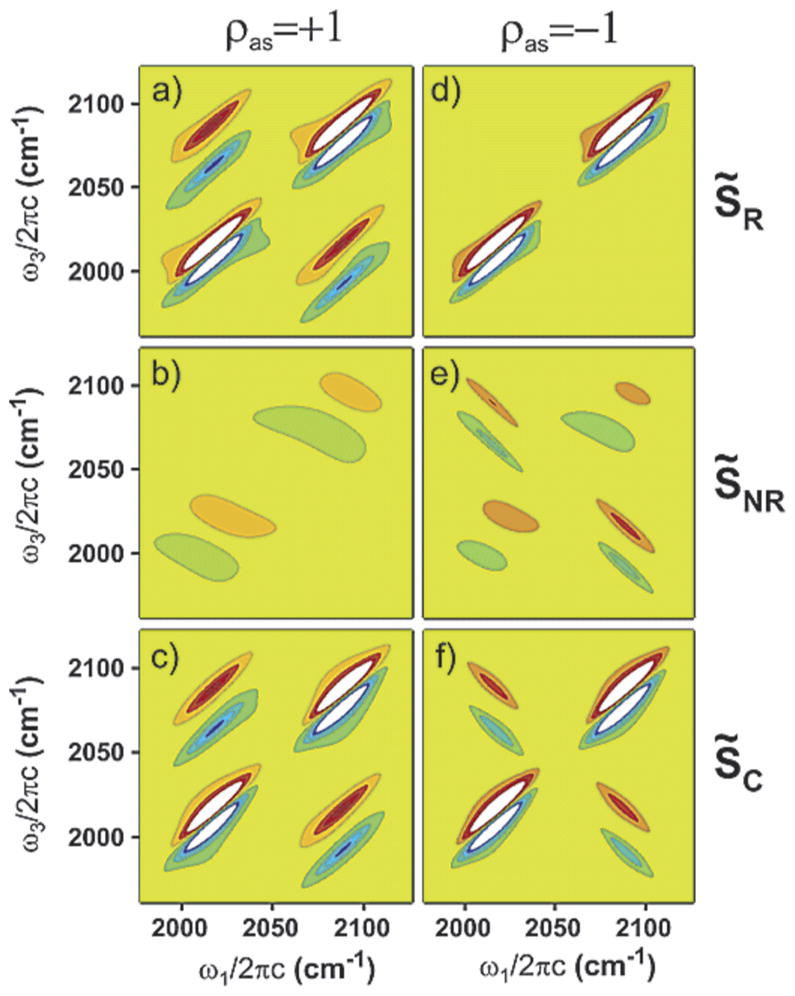
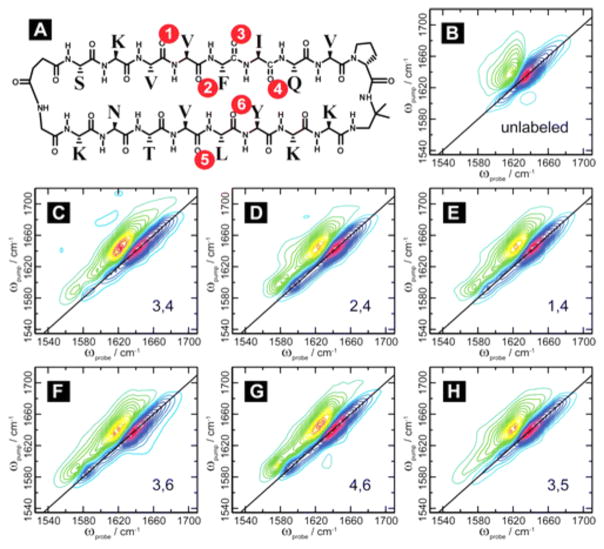
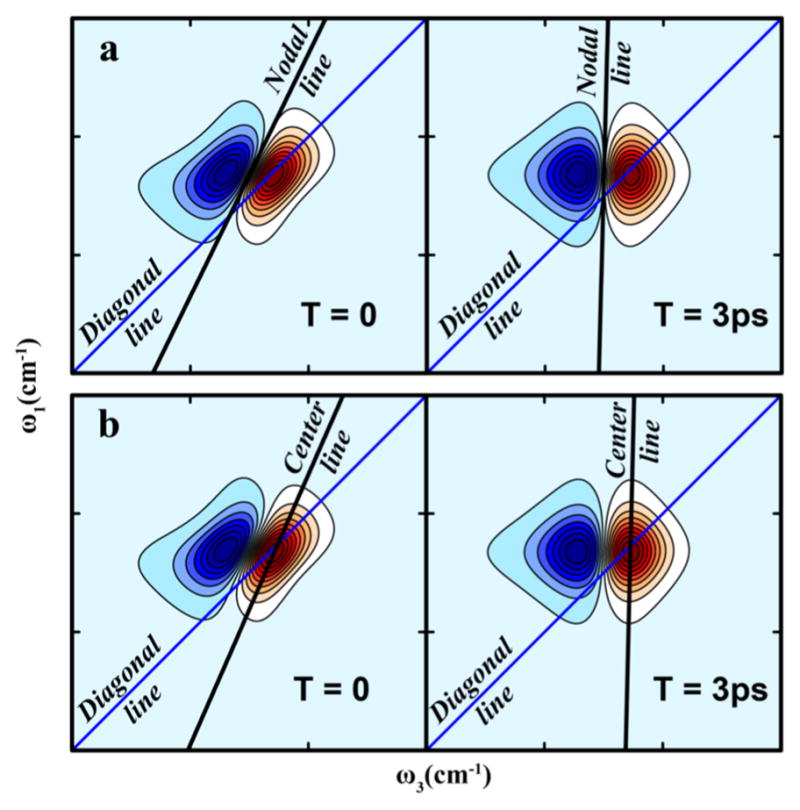
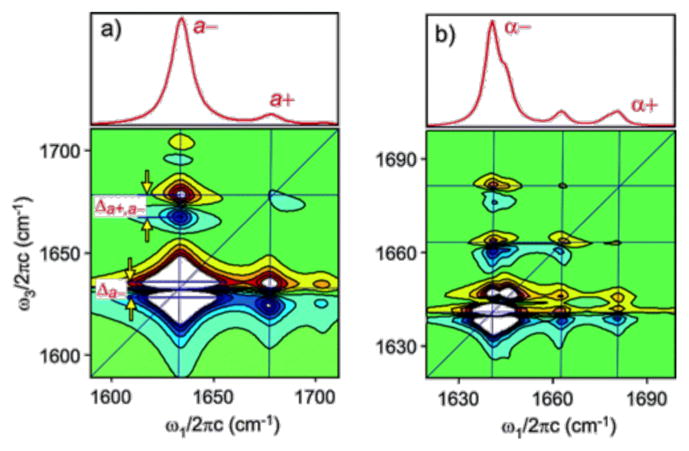
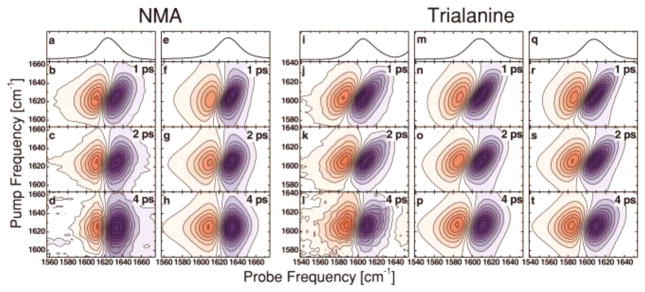

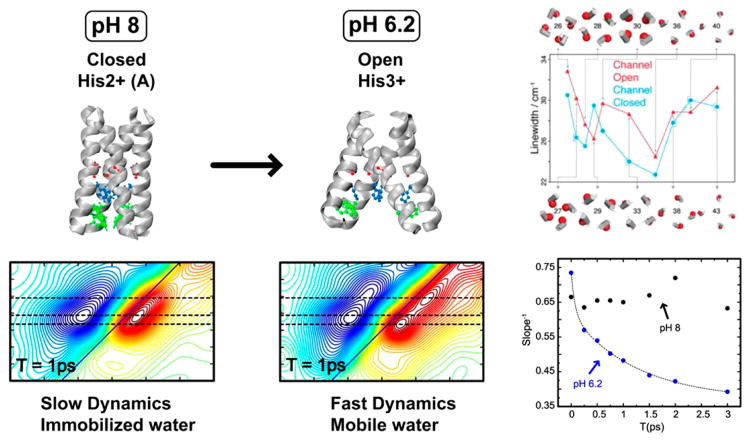
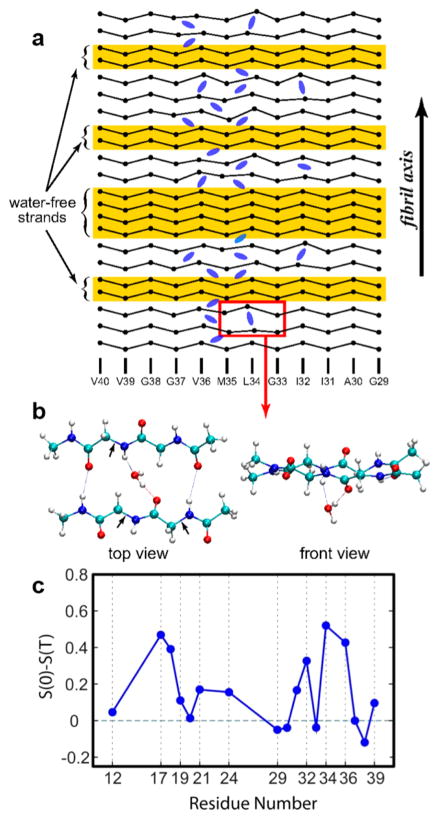
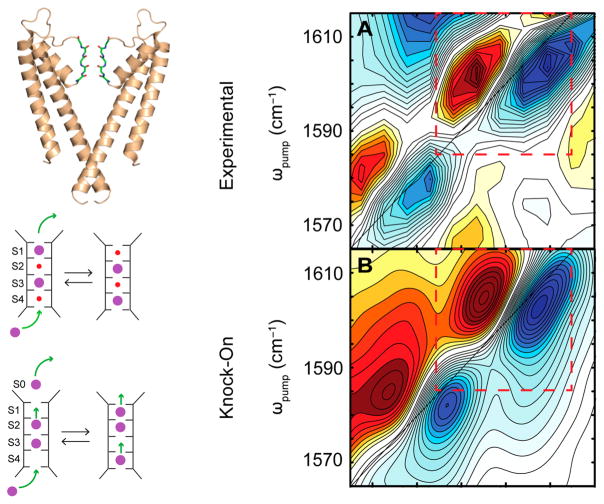
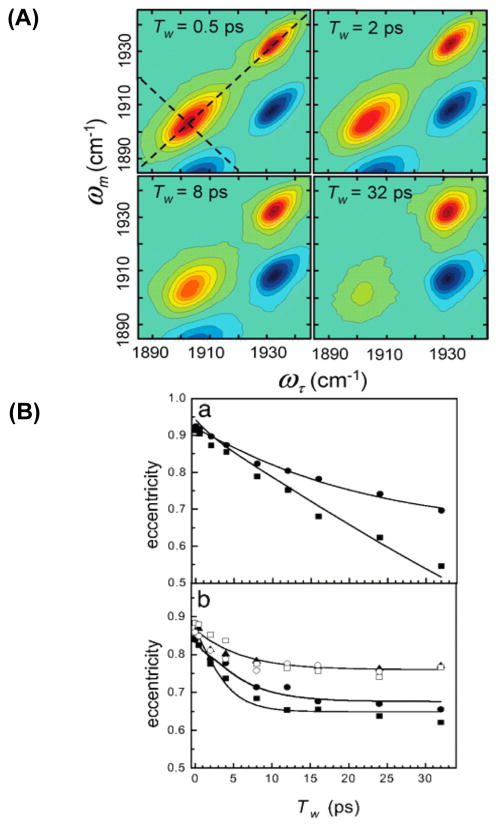
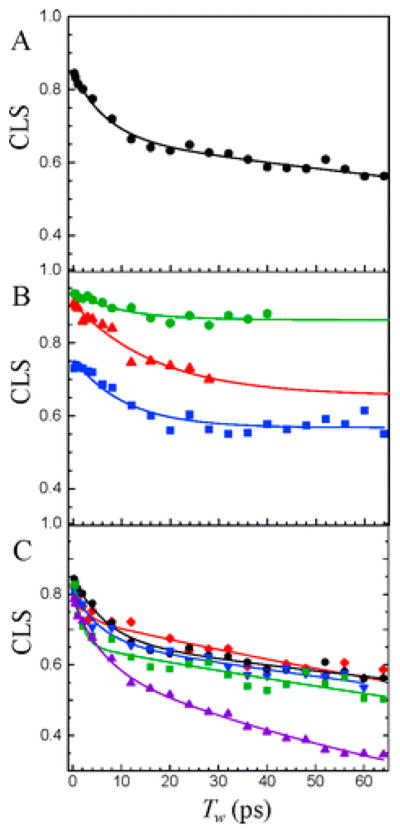
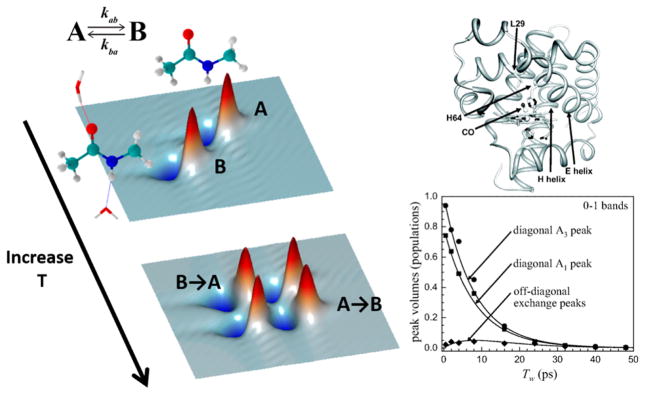
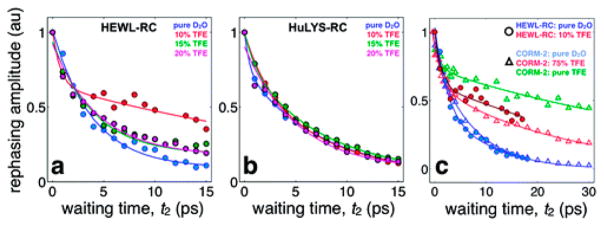
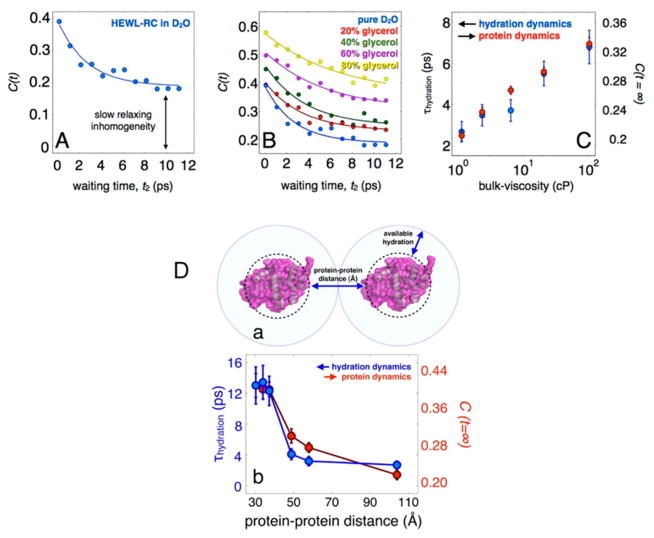
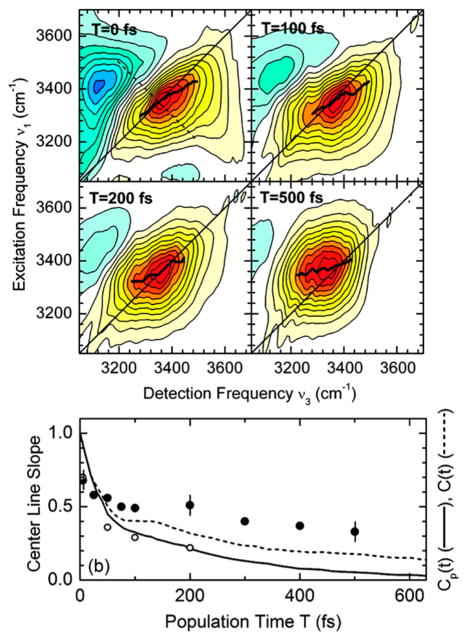
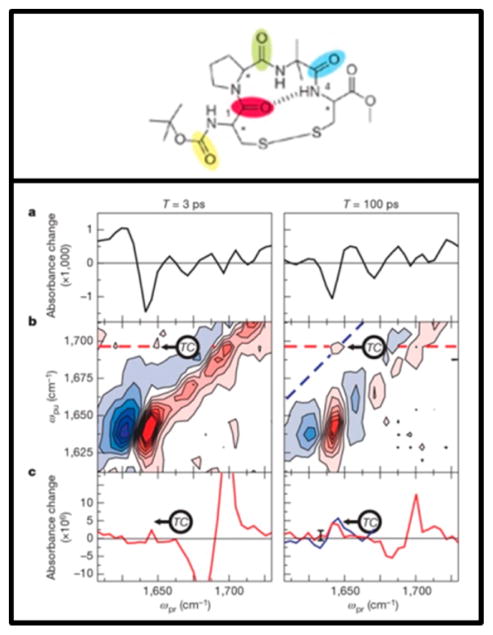
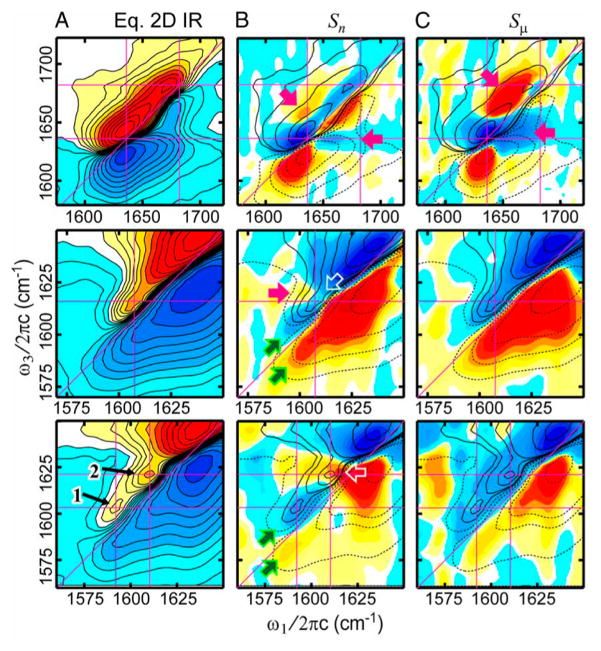
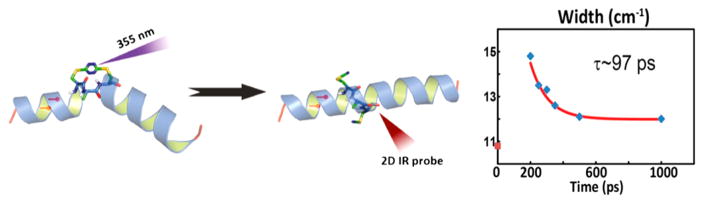
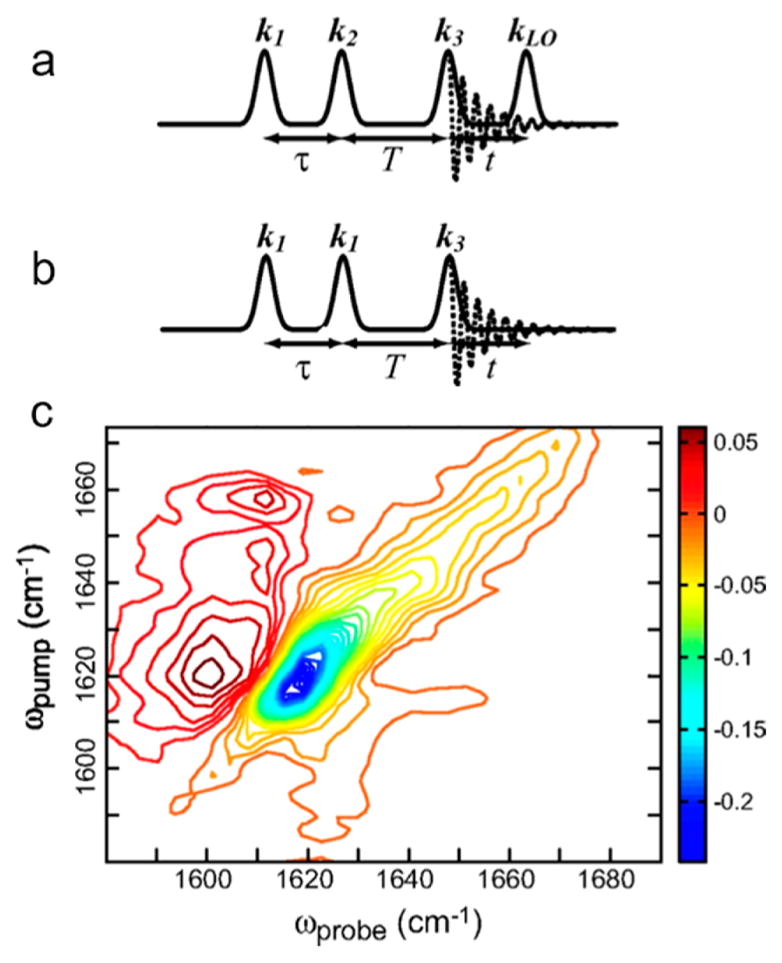
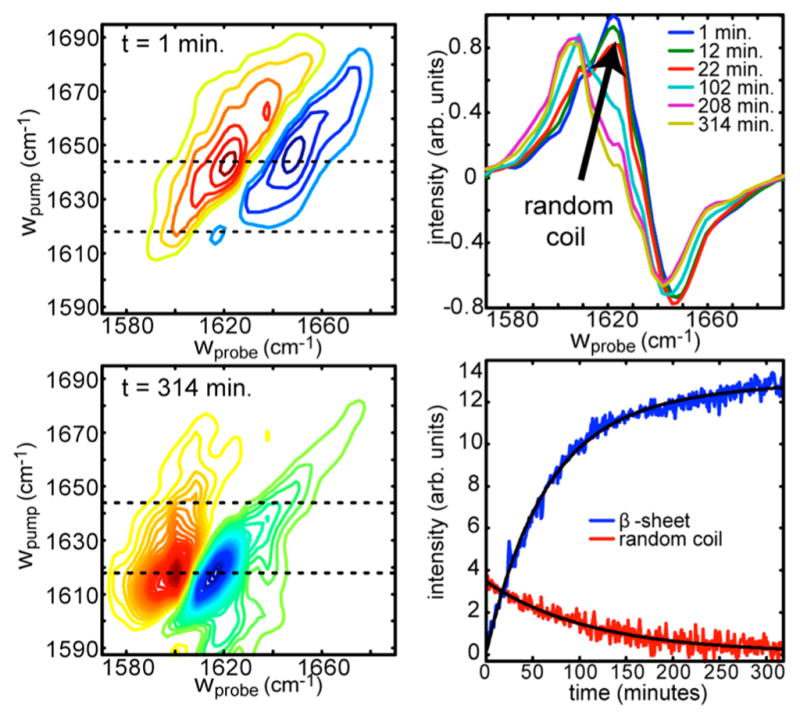
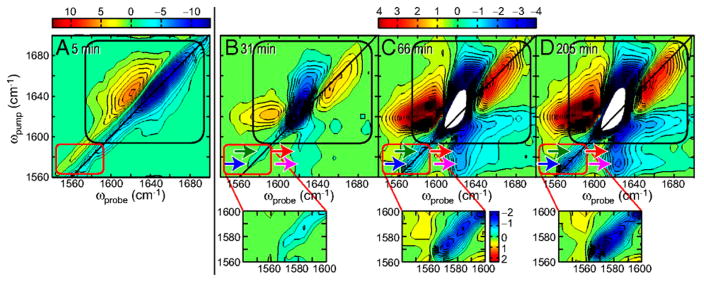
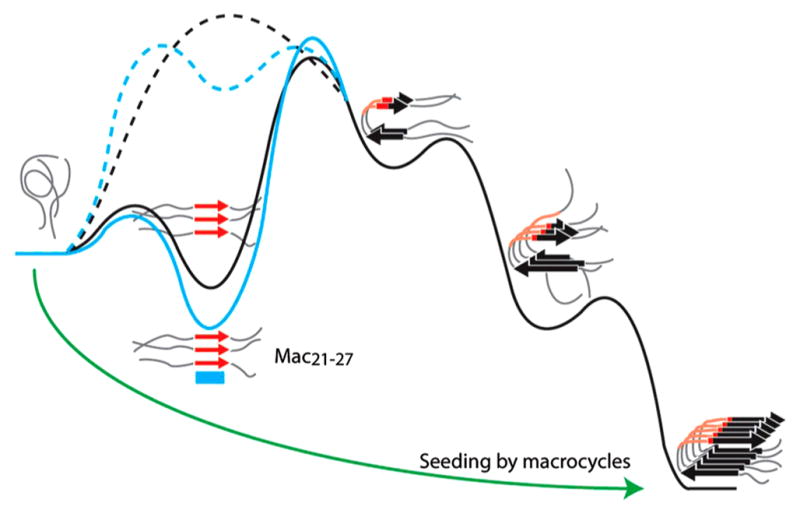
References
-
- Henzler-Wildman K, Kern D. Dynamic Personalities of Proteins. Nature. 2007;450:964–972. - PubMed
-
- Hammes-Schiffer S, Benkovic SJ. Relating Protein Motion to Catalysis. Annu Rev Biochem. 2006;75:519–541. - PubMed
-
- Fersht A. Structure and Mechanism in Protein Science: A Guide to Enzyme Catalysis and Protein Folding. 1. WH Freeman; 1998.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
